Wie das Gehirn die Seele macht
Prof.Dr.Dr.
Gerhard Roth
Direktor am Institut für Hirnforschung der Universität
Bremen
Rektor des Hanse-Wissenschaftskollegs Delmenhorst
Vortrag am 22. April
2001 bei den 51. Lindauer Psychotherapiewochen (http://www.lptw.de/)
Im Jahre 1896 verfasste der damals vierzigjährige Sigmund Freud eine umfangreiche Schrift mit dem Titel "Entwurf einer Psychologie". Sie stellte den Versuch dar, eine Theorie des "seelischen Apparates" auf der Grundlage der soeben etablierten Neuronentheorie zu entwickeln. Diese Schrift wurde nie veröffentlicht, sie erschien erst posthum (1950).
Freud hatte eine gründliche Ausbildung als Neurobiologe in einigen der damals berühmtesten Labors hinter sich. Hierzu gehörte das Labor des Physiologen und Histologen Ernst Brücke (1819 1892), eines Schülers von Johannes Müller, der als der Begründers der modernen Physiologie gilt. Neben Brücke zählten zu den Schülern von Müller Hermann von Helmholtz, Emil Dubois-Reymond und Carl Ludwig, die alle als Väter der modernen Natur- und Biowissenschaften angesehen werden. Im Labor von Brücke traf Freud auf den Physiologen Sigmund Exner und den Neurologen Josef Breuer, die ihn auf unterschiedliche Art stark beeinflussen sollten. Nach seiner Promotion in Medizin war Freud im Labor des Hirnanatomen Theodor Meynert tätig und beschäftigte sich mit der Neuroanatomie des Hirnstamms von Wirbeltieren. Meynert (1833 1892) galt neben Paul Flechsig in Leipzig als der führende europäische Neuroanatom; er versuchte psychische und psychopathologische Phänomene mehr oder weniger direkt mit der Aktivität bestimmter Hirnzentren in Verbindung zu bringen, und tat damit das, was man damals als "Gehirnmythologie" bezeichnete.
An solcher Gehirnmythologie beteiligte sich auch Freud in seinem erwähnten unveröffentlichten "Entwurf der Psychologie" von 1896, der ihm offenbar völlig zu Unrecht! lebenslang peinlich war. Man muss sich dabei vergegenwärtigen, dass erst wenige Jahre zuvor die für die heutige Neurobiologie grundlegende "Neuronen-Doktrin" veröffentlicht worden war, nämlich die Anschauung, dass das Nervensystem aus Nervenzellen, Neurone genannt, aufgebaut ist, und dass die seit langem im Gehirn beobachteten "Ganglienkugeln" und "Fasern" zusammengehören und eben die Nervenzellen mitsamt Fortsatz (Axon genannt) bilden. Völlig unklar war noch zu Freuds Studienzeiten die Frage, welche Art von Kontakten die Nervenzellen untereinander haben; der Begriff "Synapse" wurde erst einige Jahre später von Charles Sherrington geprägt. Von den elektrophysiologischen Vorgängen im Gehirn hatte man wenig, von den neurochemischen Vorgängen keinerlei Kenntnis. Hingegen wusste man über die anatomische Gliederung des menschlichen Gehirns einigermaßen Bescheid und hatte grobe Vorstellungen über die Funktionen der einzelnen Hirnzentren.
Freud hat in seinem unveröffentlichten Aufsatz vieles von dem vorweg genommen, was er einige Jahre später in der "Traumdeutung" veröffentlichte und was zur Grundlage der so genannten ersten Topik, also seiner ersten Version des Verhältnisses zwischen dem Unbewussten, dem Vorbewussten, dem Ich und dem Bewusstsein, wurde. Vom Standpunkt der Neurowissenschaften aus gesehen ist dieser Aufsatz ebenfalls interessant. Es gibt zwar einige aus heutiger Sicht völlig falsche Annahmen, vor allem das von Gustav Fechner übernommene Prinzip, alle seelischen Vorgänge seien auf Abfuhr von übermäßiger Erregung, von Spannung, angelegt, die von außen als Wahrnehmung (furcht-)erregender Umweltgeschehnisse und von innen als Triebe in die Psyche eindringen. Derart erzeugte psychische Spannung sei Unlust; Abfuhr oder zumindest Herabsetzung von Spannung erzeuge dagegen Lust. Freud konnte sich damals ebenso wenig wie alle anderen Neurobiologen vorstellen, dass es Zentren geben könne, die durch ihre Aktivität Lustgefühle erzeugen; ebenso wenig war das Prinzip der neuronalen Hemmung bekannt.
Freud war jedoch einer der ersten, der die Idee vortrug, dass das Gehirn aus untereinander verknüpften Neuronen besteht, die dadurch komplizierte Nervennetze bilden, und dass sich aus der Art der Verknüpfung der Neurone die Funktion dieser Nervennetze ergibt. Diese Vorstellung wurde aber erst durch die detaillierten Untersuchungen des spanischen Neuroanatomen Santiago Ramón y Cajal (1852 1934) zum Durchbruch verholfen; Ramón y Cajal erhielt hierfür 1906 den Nobelpreis. Freud entwickelte auch übrigens wohl unabhängig von seinem Laborkollegen Sigmund Exner (1846 1926) die Idee, dass eine gleichzeitige Tätigkeit zweier neuronaler Kontaktpunkte, die später Synapsen genannt wurde, die Leitfähigkeit dieser Kontakte erhöhte. Erst fünfzig Jahre später wurde diese Idee von Donald Hebb wieder aufgegriffen, die heute als Hebb-Synapse bekannt ist (sie müsste eigentlich Exner-Synapse heißen) und an der das Phänomen der "Langzeitpotenzierung" (LTP) als eine Grundlage von Lernen und Gedächtnisbildung studiert werden kann. Untersuchungen des amerikanischen Neurobiologen Eric Kandel zur Langzeitpotenzierung wurden im Jahre 2000 ebenfalls mit dem Nobelpreis gewürdigt.
Warum Freud seinen "Entwurf der Psychologie" nicht veröffentlichte, darüber habe ich keine genauen Auskünfte gefunden. Ich vermute, dass er die Vergeblichkeit seines Tuns, auf Grundlage der damaligen Kenntnis der Arbeitsweise des Gehirns das Psychische erklären zu wollen, einsah. Er wandte sich enttäuscht von der Neurobiologie ab und äußerte sich später an vielen Stellen skeptisch bis ablehnend über die Möglichkeiten, das Psychische neurobiologisch erklären zu können. In seinem Aufsatz "Das Unbewusste" aus dem Jahre 1915 schreibt er zur Beziehung des seelischen Apparates zur Anatomie:
"Wir wissen, dass solche Beziehungen im gröbsten existieren. Es ist ein unerschütterliches Resultat der Forschung, dass die seelische Tätigkeit an die Funktion des Gehirns gebunden ist wie an kein anderes Organ. Ein Stück weiter es ist nicht bekannt, wie weit führt die Entdeckung von der Ungleichwertigkeit der Gehirnteile und deren Sonderbeziehung zu bestimmten Körperteilen und geistigen Tätigkeiten. Aber alle Versuche, von da aus eine Lokalisation der seelischen Vorgänge zu erraten, alle Bemühungen, die Vorstellungen in Nervenzellen aufgespeichert zu denken und die Erregungen auf Nervenfasern wandern zu lassen, sind gründlich gescheitert. Dasselbe Schicksal würde einer Lehre bevorstehen, die etwa den anatomischen Ort des Systems Bw, der bewussten Seelentätigkeit, in der Hirnrinde erkennen und die unbewussten Vorgänge in die subkortikalen Hirnpartien versetzen wollte. Es klafft hier eine Lücke, deren Ausfüllung derzeit nicht möglich ist, auch nicht zu den Aufgaben der Psychologie gehört. Unsere psychische Topik hat vorläufig nichts mit der Anatomie zu tun; sie bezieht sich auf Regionen des seelischen Apparates, wo immer sie im Körper gelegen sein mögen, und nicht auf anatomische Örtlichkeiten."
Die große Enttäuschung darüber, dass die Hirnforschung seinerzeit wenig oder gar nichts zur Aufklärung des Psychischen beitragen konnten, hat Freud sein Leben lang geplagt, und er hat stets zwischen krasser Ablehnung der Neurobiologie und der Hoffnung, die Situation können sich doch ändern, geschwankt.
Dieser Standpunkt ist durchaus verständlich, denn bis zu Freuds Tod im Jahre 1939 gab es in der Hirnforschung nahezu nichts, was einen Ansatzpunkt zu einer konvergenten Entwicklung zwischen Hirnforschung und Psychotherapie bzw. Psychoanalyse hätte liefern können. Man muss dabei bedenken, dass die Mikroelektroden-Technik, mithilfe derer man die Aktivität einzelner Neurone registrieren kann, sich erst im Laufe der fünfziger Jahre des 20. Jahrhunderts entwickelte, was in den folgenden Jahrzehnten zu einer raschen Aufklärung sensorischer Funktionen, vor allem des Sehsystems, und motorischer Funktionen führte. In den siebziger und achtziger Jahren kam die Immuncytochemie hinzu, die überhaupt erst die neurochemische und neuropharmakologische Vielfalt des Gehirns enthüllte. Intrazelluläre Ableitungen und die Patch-Clamp-Technik revolutionierten unser Verständnis von den Prozessen, die bei Erregung und Hemmung an der Membran einer Nervenzelle stattfinden. In den neunziger Jahren des vergangenen Jahrhunderts kamen die bildgebenden Verfahren (Positronen-Emissions-Tomographie, PET, und funktionelle Kernspintomographie, fNMR) hinzu, die erstmals im Gegensatz zur Elektroenzephalographie (EEG) die genauere Lokalisierung der Aktivitäten des gesamten menschlichen Gehirns bei ungeöffneten Schädel ermöglichten. Durch Kombination all dieser Methoden begann erst vor rund einem Jahrzehnt die genauere Erforschung der neuronalen Grundlagen dessen, was Freud "psychischen Apparat" genannt hat.
In starker Vereinfachung kann man die Grundaussagen Freuds zu diesem "psychischen Apparat", insbesondere zum Verhältnis zwischen Bewusstem und Unbewussten und zur Rolle des Ich folgendermaßen zusammenfassen: (1) Das Unbewusste determiniert weitgehend das Bewusstsein; (2) das Unbewusste entsteht ontogenetisch weit vor dem Bewusstsein; es legt sehr früh die Grundstrukturen des Psychischen und des bewussten Erlebens fest; (3) das bewusste Ich hat keine oder nur geringe Einsicht in die unbewussten Determinanten des Erlebens und Handelns, es kann sich nicht selbst therapieren. Hieraus resultiert die Notwendigkeit der Therapie durch den Psychotherapeuten.
Bei der Frage, ob diese Grundannahmen aus Sicht der Hirnforschung gerechtfertigt sind und wie aus dieser Sicht "das Gehirn die Seele macht", geht es um folgende Teilfragen:
1. Was ist aus neurobiologischer Sicht Bewusstsein und was das Unbewusste?
2. Wo und wie im Gehirn entstehen Affekte und Emotionen?
3. Wo im Gehirn existiert das Ich und wann und wie entsteht es?
5. Was sind aus neurobiologischer Sicht psychische Erkrankungen, und wie ist Psychotherapie möglich?
Ich möchte auf jede dieser Fragen eingehen, dies kann an dieser Stelle jedoch nur andeutungsweise geschehen. Eine ausführlichere Darstellung findet sich in meinem Buch "Fühlen, Denken, Handeln Wie das Gehirn unser Verhalten steuert" (Roth, 2001).
Was ist aus neurobiologischer Sicht das Bewusste und was das Unbewusste?
Bewusstsein umfasst alle Zustände, die von einem Individuum erlebt werden, und tritt in einer Vielzahl unterschiedlicher Zustände auf. Hierzu gehören a) Sinneswahrnehmungen von Vorgängen in der Umwelt und im eigenen Körper, b) mentale Zustände und Tätigkeiten wie Denken, Vorstellen und Erinnern, c) Emotionen, Affekte, Bedürfniszustände, d) Erleben der eigenen Identität und Kontinuität, e) "Meinigkeit" des eigenen Körpers, f) Autorschaft und Kontrolle der eigenen Handlungen und mentalen Akte, g) Verortung des Selbst und des Körpers in Raum und Zeit, h) Realitätscharakter von Erlebtem und Unterscheidung zwischen Realität und Vorstellung.
Bewusstseinszustände treten entweder als Hintergrundbewusstsein auf, welches Ich-Identität, "Meinigkeit" und willentliche Kontrolle des Körpers, Verortung des Ich und des Körpers in Raum und Zeit und den Realitätscharakter des Erlebten betrifft, oder als schnell wechselndes Aktualbewusstsein, welches sich aus den jeweiligen Sinneserlebnissen, den Emotionen, den kognitiven (Denken, Vorstellen, Erinnern) und exekutiven Zuständen (Handlungsplanung und Handlungskontrolle) zusammensetzt.
Geschehnisse können grundsätzlich nur dann bewusst werden, wenn sie mit der Aktivität der assoziativen Großhirnrinde verbunden sind (Roth, 1996, 2001). Allerdings sind nur wenige Prozesse, die im assoziativen Cortex ablaufen, aktuell bewusst. Zum assoziativen Cortex gehören der hintere Scheitellappen (posteriorer parietaler Cortex), der Schläfenlappen (temporaler Cortex) und sein Übergang zum Hinterhauptslappen (okzipitaler Cortex) und schließlich der Stirnlappen (präfrontaler und orbitofrontaler Cortex). An der Entstehung und Kontrolle solcher Bewusstseinszustände sind jedoch zahlreiche andere Hirngebiete beteiligt, deren Aktivität selbst grundsätzlich nicht bewusstseinsfähig ist. Hierzu gehören isocorticale (d. h. sechsschichtige) sensorische und motorische Hirnrindengebiete, allocorticale (d.h. drei- bis fünfschichtige) Hirnrindengebiete (cingulärer, entorhinaler und insulärer Cortex, Hippocampus) sowie subcorticale Zentren, die ich noch schildern werde.
Vor dem Hintergrund dieser Beschreibung bewusster Zustände umfasst aus Sicht der Hirnforschung und der experimentellen Psychologie das Unbewusste folgende Inhalte: (1) Inhalte, die einmal bewusst waren, aber ins Unbewusste abgesunken sind und unter günstigen Bedingungen wieder bewusst gemacht ("erinnert") werden können (z. B. nicht-aktivierte Inhalte des deklarativen Gedächtnisses); (2) vorbewusste Inhalte von Wahrnehmungsvorgängen, die nach hinreichender Aktivierung der assoziativen Großhirnrinde bewusst werden (die geschieht mit einer Verzögerung von durchschnittlich 300 bis 500 Millisekunden nach Reizbeginn); (3) unterschwellige (subliminale) Wahrnehmungen; (4) Vorgänge in Gehirnregionen außerhalb der assoziativen Großhirnrinde, die grundsätzlich unbewusst ablaufen; (5) alle perzeptiven, kognitiven und emotionalen Prozesse, die im Gehirn des Fötus, des Säuglings und des Kleinkindes vor Ausreifung des assoziativen Cortex ablaufen. Man nimmt an, dass sich beim Menschen Ich-bezogene Bewusstseins- und Gedächtnisinhalte erst ab Ende des dritten Lebensjahres entwickeln; dies würde die Idee Freuds von einer infantilen Amnesie bestätigen.
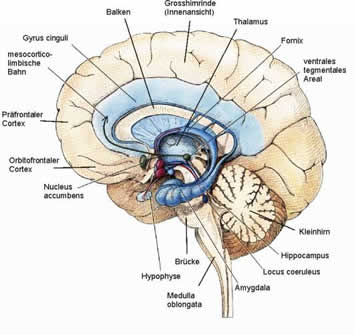
Aus Sicht der Hirnforschung und der Neuropsychologie besteht kein Zweifel daran, dass die soeben beschriebenen unbewussten Gehirnvorgänge höchst wirksam sind und bewusste Vorgänge beeinflussen (vgl. Goschke, 1996, 1997). Neben den subliminalen und vorbewussten Komponenten der Wahrnehmung sind dies vor allem Vorgänge im limbischen System. Das limbische System durchzieht das gesamte Gehirn; seine Tätigkeit erleben wir als Affekte und Emotionen bzw. affektive und emotionale Einfärbungen von Wahrnehmung, Vorstellung, Erinnerung und Handlungsplanung (Abbildung 1).
Abbildung 1: Medianansicht des menschlichen Gehirns mit den wichtigsten limbischen Zentren. Diese Zentren sind Orte der Entstehung von positiven (Nucleus accumbens, ventrales tegmentales Areal), und negativen Gefühlen (Amygdala), der Gedächtnisorganisation (Hippocampus), der Aufmerksamkeits- und Bewusstseinssteuerung (basales Vorderhirn, Locus coeruleus, Thalamus) und der vegetativen Funktionen (Hypothalamus). (Aus Roth, 2001, nach Spektrum/Scientific American, 1994, verändert).
Die wichtigsten limbischen Zentren sind (1) ventrales tegmentales Areal (VTA) und zentrales Höhlengrau (PAG) im ventralen Mittelhirn (Tegmentum des Mesencephalon); (2) Hypothalamus, ventrales Pallidum, Mammillarkörper, anteriore, mediale, intralaminare und Mittellinienkerne des Thalamus im Zwischenhirn (Diencephalon); (3) orbitofrontaler, inferiorer temporaler, cingulärer, entorhinaler und insulärer Cortex, Amygdala, Septum, ventrales Striatum/Nucleus accumbens im Endhirn (Telencephalon). Im folgenden werden diese Zentren entsprechend ihrer funktionalen Rolle besprochen.
Der Hypothalamus ist das grundlegende Kontrollzentrum für biologische Grundfunktionen wie Nahrungs- und Flüssigkeitsaufnahme, Sexualverhalten, Schlaf- und Wachzustand, Temperatur- und Kreislaufregulation, Angriffs- und Verteidigungsverhalten und für die damit verbundenen "angeborenen" Trieb- und Affektzustände. Entsprechend seinen Funktionen ist der Hypothalamus mit nahezu allen anderen Teilen des Gehirns verbunden. Er hat enge Beziehungen zu den limbischen Anteilen des Telencephalon, d.h. zum Septum, zur Amygdala, zum Hippocampus und zum orbitofrontalen Cortex. Er kontrolliert die Hirnanhangdrüse (Hypophyse) und hierüber das Hormonsystem (besonders auch die hormonale Stressreaktion), die vegetativ-autonomen Kerngebiete des Hirnstamms und Rückenmarks, die ihrerseits Ausgangspunkt des sympathischen und parasympathischen Nervensystems sind.
Das Zentrale Höhlengrau (periaquäduktales Grau, PAG) ist neben dem Hypothalamus und dem Zentralkern der Amygdala (s. unten) das wichtigste Zentrum für angeborene affektive Zustände und Verhaltensweisen. Es kann unabhängig von höheren affektiv-emotionalen Zentren wie der basolateralen Amygdala, dem Hippocampus oder der Großhirnrinde arbeiten, wird aber meist von diesen beeinflusst. Es kontrolliert wie der Hypothalamus Sexualverhalten, Aggression, Verteidigung und Beutefang/Nahrungsaufnahme. Eine wichtige Rolle spielt es auch bei der unbewussten Schmerzreaktion und affektiv-emotionalen Vokalisationen (Schmerzschreie, Stöhnen, Klagen usw.).
Die Amygdala nimmt anatomisch wie funktional die entscheidende Rolle bei der Produktion und Steuerung von Emotionen ein. Sie wird als das Zentrum der furcht- und angstgeleiteten Verhaltensbewertung angesehen; Läsionen der Amygdala führen zum Fortfall der Furcht- oder Angstkomponente beim Erleben von Geschehnissen.
Die Amygdala ist anatomisch und funktional sehr unterschiedlich zusammengesetzt. Sie besteht aus einer corticomedialen Kerngruppe, die vornehmlich mit der Verarbeitung olfaktorischer Informationen (vor allem von Pheromonen) zu tun hat, einer basolateralen Kerngruppe (bei Primaten einschließlich Mensch besonders stark entwickelt), die mit Furchtkonditionierung zu tun hat, und dem bereits genannten Zentralkern, der den Ausgangsbereich der Amygdala bildet und wie der Hypothalamus "angeborene" affektive Funktionen besitzt. Die Amygdala unterhält direkte oder über den mediodorsalen Thalamus ziehende rückläufige Verbindungen mit dem assoziativen Cortex, und zwar vornehmlich mit dem orbitofrontalen, temporalen und cingulären Cortex. Allgemein sind die von der Amygdala zum Isocortex verlaufenden Bahnen stärker als die Bahnen in umgekehrter Richtung. Direkte sensorische (somatosensorische, visuelle und auditorische) Eingänge erhält die Amygdala über den Thalamus, die im basolateralen Kernbereich enden. Hier enden auch Eingänge vom Hippocampus und der umgebenden Rinde, die kontextuelle Gedächtnisinhalte liefern (siehe unten).
Direkt bzw. mittelbar über den Hypothalamus und das PAG wirkt die Amygdala auf das gesamte hormonale und vegetative System ein, z.B. durch Aktivierung des sympathischen und parasympathischen Systems (vegetative Reaktionen) und durch Aktivierung des dopaminergen, noradrenergen und cholinergen Systems (Erhöhung des Wachheitszustandes und der Verhaltensbereitschaft), der Kreislauf- und Atemfunktionen, der Gesichtsmimik, der Verteidigungs- und Fluchtreaktionen und der Ausschüttung von Corticosteroiden bei der Stressreaktion (über Hypothalamus, Hypophyse und Nebennierenrinde).
Die Amygdala stellt zugleich das Verbindungszentrum zwischen erlernter (d.h. konditionierter) und angeborener Furcht einerseits und den damit verbundenen autonom-vegetativen Reaktionen andererseits dar, indem in ihr sensorische Informationen (über den Thalamus einlaufend) und kontextuelle Gedächtnisinhalte (über die Hippocampusformation) über bestimmte negative Ereignisse mit den genannten angeborenen vegetativen und affektiven Furchtreaktionen verbunden werden. Von einer Reihe von Autoren wird die Beteiligung der Amygdala auch an nicht furchtbedingten oder gar positiv besetzten, appetitiven Zuständen beim Lernen angenommen, insbesondere im Zusammenhang mit der Nahrungsaufnahme (Rolls, 1999).
Der Hippocampus ist der Organisator des bewusstseinsfähigen, deklarativen Gedächtnisses, und zwar zusammen mit der ihn umgebenden Rinde, die aus dem entorhinalen, perirhinalen und parahippocampalen Cortex (zusammen EPPC genannt) besteht. Eingänge aus dem assoziativen Isocortex laufen über den perirhinalen bzw. parahippocampalen Cortex und von dort über den entorhinalen Cortex, der das corticale "Eingangstor" zum Hippocampus bildet. Direkte subcorticale Afferenzen des Hippocampus kommen vom basalen Vorderhirn (genauer vom medialen Septum), von der basolateralen Amygdala, vom Hypothalamus, von den limbischen Thalamuskernen, den Raphe-Kernen, dem Locus coeruleus und dem PAG (um nur die wichtigsten Zentren zu nennen). Die Ausgänge des Hippocampus ziehen über den EPPC wieder zu isocorticalen Zielgebieten zurück sowie zur Amygdala, zum basalen Vorderhirn, zum Nucleus accumbens und zum Hypothalamus.
Die Speicherung des Wissens findet nicht im Hippocampus und im EPPC selbst statt, sondern modalitäts- und funktionsspezifisch in den verschiedenen Rindenarealen. Entsprechend befindet sich das visuelle Gedächtnis in den visuellen Cortexregionen, das auditorische Gedächtnis in den auditorischen Arealen, die sprachlichen Erinnerungen in den Sprachzentren usw. Eine bilaterale Zerstörung des Hippocampus führt zu zeitlich begrenzter retrograder Amnesie, d.h. zum Verlust von Teilen des Altgedächtnisses, sowie zur anterograden Amnesie, d.h. zur Unfähigkeit, neue Inhalte in das deklarative bzw. semantische und episodische Gedächtnis einzufügen. Für derartige Patienten ist alles neu, was ihnen nicht seit langem bekannt und "eingeschliffen ist.
Neuere Untersuchungen (Tulving und Markowitsch, 1998; Markowitsch, 1999; Aggleton und Brown, 1999) unterscheiden innerhalb des deklarativen Gedächtnisses ein episodisches Gedächtnis, das sich immer auf inhaltlich, räumlich und zeitlich konkrete Erlebnisse mit Bezug auf die eigene Person bezieht ("was mir Montag voriger Woche in Hamburg passierte"), und ein Wissensgedächtnis, das sich auf personen-, orts- und zeitunabhängige Tatsachen bezieht ("der Mars ist ein Planet"). Die Einspeicherung des episodischen Gedächtnisses wird dem Hippocampus im engeren Sinne zugeordnet, das Wissensgedächtnis dem EPPC.
Hippocampus, EPPC und Amygdala arbeiten im Bereich des deklarativen und emotionalen Gedächtnisses "arbeitsteilig". Im Rahmen einer klassischen Konditionierung, in dem bei Normalpersonen ein Nebelhorn Schreckreaktionen auslöste, konnten Patienten mit einer bilateralen Schädigung der Amygdala genau angeben, welcher sensorische Stimulus mit dem Schreckreiz gepaart worden war, sie zeigten aber keinerlei vegetative Furchtreaktion, gemessen über die Veränderung des Hautwiderstands (Bechara et al., 1995). Sie entwickelten also keine Furcht- oder Schreckempfindungen und nahmen die Ereignisse "emotionslos hin. Umgekehrt hatten Patienten mit bilateraler Schädigung des Hippocampus keine bewusste Information über die Paarung von sensorischem Reiz und Schreckreiz, zeigten aber eine deutliche vegetative Furchtreaktion. Während also ihr emotionales Gedächtnis funktionierte, versagte ihr deklaratives Gedächtnis, was nach der Hippocampus-Läsion auch zu erwarten war. Die Patienten mit Amygdala und ohne Hippocampus erlebten also Angst und Schrecken, ohne zu wissen, warum.
Ein "Gegenspieler" der Amygdala ist das mesolimbische System, das aus dem ventralen tegmentalen Areal (VTA), dem lateralen Hypothalamus, dem Nucleus accumbens/ventralen Striatum und dem ventralen Pallidum besteht. Dieses System ist durch den Neuromodulator Dopamin charakterisiert. Es dominiert bei der Registrierung und Verarbeitung natürlicher Belohnungsereignisse und stellt offenbar das zerebrale Belohnungssystem oder zumindest einen wichtigen Teil dieses Systems dar. Intracraniale Selbststimulation im mesolimbischen System führt bei Ratten und Menschen zu stark lustbetonten Zuständen, allerdings nur dann, wenn sich der Organismus in einem Bedürfniszustand befindet (Panksepp, 1998).
Das mesolimbische System stellt auch den Wirkort für Drogen einschließlich Alkohol und Psychopharmaka dar. Letztere erhöhen direkt oder über die Hemmung hemmender Interneurone den Dopaminspiegel im VTA bzw. im Nucleus accumbens. Allerdings scheint nach neueren Erkenntnissen eine Erhöhung des Dopaminspiegels nur für die Suchtentstehung, nicht aber für die Aufrechterhaltung der Sucht notwendig zu sein. Dopamin scheint im Lichte dieser Befunde eher als Signal für die Assoziation von Belohnung und bestimmten Ereignissen (und damit als Voraussage von Belohnung) denn als "Belohnungsstoff" selbst zu fungieren. Diese Funktion kommt offenbar eher den hirneigenen Opiaten zu, deren Wirkort ebenfalls vornehmlich das mesolimbische System ist (Panksepp, 1998; Spanagel und Weiss, 1999).
Der cinguläre Cortex ist bei Aufmerksamkeitsleistungen stets zusammen mit dem dorsolateralen präfrontalen und dem posterioren parietalen Cortex aktiv. Er spielt bei der emotionalen Schmerzwahrnehmung eine wichtige Rolle; hierbei steht er in enger Beziehung mit anderen Schmerzzentren, nämlich dem insulären Cortex, den medialen Thalamuskernen und dem zentralen Höhlengrau. Nach Zerstörung der cingulären Rinde "vergessen Versuchstiere die zuvor gelernten schmerzbedingten Abwehr- und Vermeidungsreaktionen oder die negative Gefühlskomponente des Schmerzes.
Der präfrontale Cortex (PFC) im weiteren Sinne wird eingeteilt in einen dorsolateralen und einen orbitofrontalen, d.h. über den Augenhöhlen (Orbita) liegenden Teil. Er ist verbunden mit den bewussten Komponenten des Ich-Gefühls und Charakters. Der dorsolaterale PFC erhält in seinem dorsalen Teil corticale Eingänge vornehmlich aus dem hinteren Parietallappen und hat mit Bewegungen und räumlicher Strukturierung von Sinneswahrnehmungen zu tun sowie mit räumlicher Aufmerksamkeit (in Zusammenarbeit mit dem hinteren parietalen Cortex und dem cingulären Cortex). In seinem lateralen Teil erhält er Eingänge vornehmlich aus dem Temporallappen und befasst sich mit Objektwahrnehmung, dem Einschätzen gegenstands- und situationsbezogener Geschehnisse, mit kontextgerechtem Handeln und Sprechen und mit der Entwicklung von Zielvorstellungen (Petrides und Pandya, 1999; Davidson und Irwin, 1999). Der dorsolaterale PFC ist Sitz des Arbeitsgedächtnisses, d.h. der Fähigkeit, wichtige Wahrnehmungsinhalte für einige Sekunden im Bewusstsein zu halten und sie geistig zu verarbeiten (Petrides, 2000). Läsionen des dorsolateralen PFC führen auch zur Unfähigkeit, die sachliche Relevanz externer Ereignisse einzuschätzen.
Der orbitofrontale Cortex (OFC) erhält seine wichtigsten Eingänge von allocorticalen und subcorticalen limbischen Zentren, d.h. vom Gyrus cinguli, vom EPPC, von der Amygdala und vom mesolimbischen System, und befasst sich mit den motivationalen und emotionalen Aspekten von Situationen und Handlungen. Patienten mit Läsionen im orbitofrontalen Cortex sind unfähig, positive oder negative Konsequenzen ihrer Handlungen vorauszusehen, wenngleich unmittelbare Belohnung oder Bestrafung von Aktionen ihr weiteres Handeln beeinflussen können (Bechara et al., 1997; Davidson und Irwin, 1999). Sie gehen wider besseres Wissen Risiken ein. Ebenso zeigen sie einen Hang zur "Perseveration", also zum hartnäckigen Verharren bei einer Sache, und einen Verlust der Verhaltensspontaneität und Kreativität. Derartige Läsionen führen auch zum Verlust der Fähigkeit, den sozial-kommunikativen Kontext zu erfassen, z.B. die Bedeutung von Szenendarstellungen oder die Mimik von Gesichtern,. Allgemein verflacht die Persönlichkeit, der Patient wird völlig "unemotional" (Damasio, 1994).
Der orbitofrontale Cortex ist auch der "Sitz" ethischer und moralischer Vorstellungen. Patienten mit Schädigung des orbitofrontalen Cortex in frühester Jugend zeigen ein schwer asoziales Verhalten, sie sind unerziehbar und unbelehrbar (Anderson et al., 1999). Sie haben bei ihrem Verhalten auch keinerlei Gewissensbisse und zeigen keinerlei Einsicht in ihr Verhalten. Die Tatsache, dass sie in normaler Umgebung aufwuchsen, hatte keinerlei positive Wirkung auf ihr Verhalten. Im Fall der Schädigung des OFC im Erwachsenenalter besteht nach Auffassung der Autoren die Schwierigkeit der Betroffenen in der Umsetzung von Erfahrung in sozial angepasstes Verhalten, über das diese Patienten durchaus verfügen. Im Fall der Schädigung des OFC in frühester Jugend wird das Sammeln solcher Erfahrung völlig verhindert; es gibt also nichts, worauf sie in Entscheidungssituationen an unbewusster, impliziter "moralischer Anweisung" hätten zurückgreifen können.
Mit dem limbischen System eng verbunden sind die neuromodulatorischen Systeme. Ihre Ausgangsneurone produzieren die Neuromodulatoren Noradrenalin, Serotonin, Dopamin und Acetylcholin und verteilen diese im Gehirn. Im Gegensatz zu den Transmittern Glutamat, Gamma-Aminobuttersäure und Glycin, welche die "schnelle", d.h. im Millisekundenbereich ablaufende synaptische Übertragung vermitteln, arbeiten Neuromodulatoren langsamer (d.h. im Bereich von Sekunden) und beeinflussen über intrazelluläre Signalketten die Wirkung der "schnellen" Transmitter.
Das erste der neuromodulatorischen Systeme ist das noradrenerge, d.h. durch Noradrenalin charakterisierte System. Es hat seinen Ausgangsort im Locus coeruleus ("blauer Kern") der lateralen retikulären Formation und sendet über weitreichende Fasern Noradrenalin in alle Teile des limbischen Systems und des assoziativen Cortex. Es vermittelt Erregung und unspezifische Aufmerksamkeit und stellt eine Komponente des Stress-Systems dar. Das zweite ist das serotonerge, d.h. durch Serotonin gekennzeichnete System. Es geht von dem an der Mittellinie ("Raphe") des Hirnstamms gelegenen Raphe-Kern aus und übt auf limbisches System und Cortex einen dämpfenden, beruhigenden und Wohlbefinden auslösenden Effekt aus. Das dritte ist das dopaminerge System, das vom ventralen tegmentalen Areal im Mittelhirn und dem Nucleus accumbens im Endhirn ausgeht. Es schickt über seine Fasern Dopamin vornehmlich in den präfrontalen und orbitofrontalen Cortex (ein weiterer, für die Bewegungssteuerung wichtiger Teil besteht im dopaminergen Projektionssystem zwischen Substantia nigra und Striatum). Es wirkt antreibend und belohnend bzw. belohnungs-versprechend (s. oben zum mesolimbischen System). Das vierte ist das cholinerge System, das im basalen Vorderhirn lokalisiert ist und Acetylcholin in den Hippocampus, die Amygdala und den assoziativen Cortex aussendet. Es vermittelt gezielte ("fokussierte") Aufmerksamkeit und übt damit einen wichtigen Einfluss auf das Gedächtnis aus.
Diese neuromodulatorischen Systeme beeinflussen zusammen mit einer großen Anzahl von Neuropeptiden unseren emotionalen und psychischen Zustand, indem sie die Veränderung der synaptischen Kopplungen von Netzwerken in der Großhirnrinde und in subcorticalen limbischen Zentren induzieren und kontrollieren, wobei sie selbst von diesen limbischen Zentren kontrolliert werden. "Oberste" Kontrolleure sind dabei der Hypothalamus, die Amygdala und der Hippocampus, d.h. die Zentren für Affekte, Emotionen und Gedächtnis.
"Ich" ist ähnlich wie "Geist" und "Bewusstsein" einer der komplexesten Begriffe der Geistesgeschichte, und diese Komplexität kann hier nicht weiter referiert werden (s. hierzu Newen und Vogeley, 2000). Ich werde im Folgenden ausschließlich die psychologischen und neurobiologischen Aspekte des Ich berücksichtigen.
In Entsprechung zu den oben genannten Bewusstseinszuständen ist das Ich modular, d.h. aus funktional unterschiedlichen Untereinheiten aufgebaut. Hierzu gehören:
(1) das Körper-Ich (dies ist mein Körper),
(2) das Verortungs-Ich (ich befinde mich gerade an dem und dem Ort),
(3) das Ich als Zentrum individuellen Verhaltens und Erlebens (perspektivisches Ich),
(4) das Ich als Subjekt perzeptiver, kognitiver und emotionaler Leistungen und Zustände (ich habe diese Wahrnehmungen, Ideen, Gefühle),
(5) das Handlungs-Ich (ich tue gerade das und das),
(6) das Autorschafts- bzw. Zurechnungs-Ich (ich bin Verursacher und Kontrolleur meiner Gedanken und Handlungen),
(7) das autobiographische Ich (ich bin derjenige, der ich gestern/früher war),
(8) das sprachliche Ich (Reden über sich selbst als überdauernde Einheit),
(9) das (selbst-)reflexive Ich (Nachdenken über sich selbst), und
(10) das ethische Ich bzw. das Gewissen.
Man unterscheidet diese verschiedenen Ich- und Bewusstseinszustände vor allem deshalb, weil sie "dissoziieren", d. h. unabhängig voneinander beeinträchtigt sein können. Es gibt entsprechend Patienten, die ein normales Ich-Bewusstsein besitzen, aber nicht wissen, wer sie sind; andere wiederum behaupten, der sie umgebende Körper bzw. einzelne Körperteile gehörten nicht zu ihnen (vgl. Sacks, 1987; Lurija, 1991). Auch kann eine Leugnung der Autorschaft eigener Ideen und Handlungen auftreten, und zwar aufgrund neurologischer oder psychischer Erkrankungen ("Ich werde gedacht" usw.).
Beeinträchtigungen und Verletzungen des assoziativen Cortex im Bereich des Scheitel-, Schläfen- und Frontallappens führen zu tiefgreifenden Ich-Störungen. Störungen im rechten Parietallappen führen zur Beeinträchtigung des Körper-Ich, der Verortung des Selbst im Raum, zu Egozentrismus und Verlust der Fähigkeit, die Perspektive zu wechseln. Störungen im rechten unteren und medialen Temporallappen führen zu Pedanterie in Sprache und Handeln, zu Egozentrik und verstärkter Aggressivität. Man spricht in diesem Zusammenhang von einer "Temporallappen-Persönlichkeit". Beeinträchtigung im medialen Temporallappenbereich (Amygdala) führt zu erhöhten Angstzuständen, Verletzung des Bereichs der Hippocampus-Formation und der entorhinalen Rinde zu anterograder und retrograder Amnesie, Verletzung des anterioren Pols des Temporallappens zum Verlust des autobiographischen Gedächtnis. Beeinträchtigungen im Bereich des dorsalen präfrontalen Cortex führen zum Verlust divergierenden Denkens, zur Unfähigkeit zur Entwicklung alternativer Strategien und spontanen Handelns, zu unflexiblem Handeln, zu Fehleinschätzung der Relevanz externer Ereignisse. Beeinträchtigung im Bereich des ventralen (orbitofrontalen) Cortex führen zu erhöhter Risikobereitschaft, allgemein zu "asozialem" Verhalten, d. h. Verletzung von gesellschaftlichen, ethischen, moralischen Regeln.
Diese Befunde unterstreichen, dass das Ich ein Gebilde ist, das aus vielfältigen, z.T. völlig unterschiedlichen Komponenten zusammengesetzt ist, die sich auf noch unbekannte Weise in jeder Sekunde unseres Selbsterlebens in unterschiedlicher Weise zusammenfügen. Dies erklärt auch das typische Fluktuieren des Ich-Gefühls. Die unterschiedlichen Ich-Zustände treten parallel zur Ausreifung des Gehirns zu unterschiedlichen Zeiten der kindlichen Entwicklung auf (Details in Roth, 2001). Gleichzeitig bekräftigen diese Befunde die Annahme Freuds, dass das Ich bewusste und unbewusste Anteile hat, denn die cortical-bewussten Ich-Zustände kommen wie geschildert nur aufgrund des Zusammenwirkens mit den unbewussten limbischen Zentren zustande.
Das Über-Ich Freuds lässt sich ohne große Schwierigkeiten im orbitofrontalen Cortex ansiedeln. Wie bereits geschildert, sind hier die in der Kindheit und Jugend erworbenen moralischen und ethischen Regeln niedergelegt. Es ist in diesem Zusammenhang höchst bemerkenswert, dass der orbitofrontale Cortex derjenige Hirnteil ist, der am spätesten, d.h. erst zum Ende der Pubertät, ausreift, d.h. zu einem Zeitpunkt, an dem die (meisten) Jugendlichen sprichwörtlich "zur Vernunft" kommen.
Das Ich ist in seinen vielfältigen Ausprägungen also ein ontogenetisch spätes Produkt des Gehirns. Als autobiographisches, sprachlich vermitteltes und reflexives Ich bildet es sich nicht vor dem Ende des dritten Lebensjahres aus. Im Gegensatz hierzu beginnt das limbische System seine Arbeit bereits im Mutterleib und setzt sie verstärkt in den ersten Wochen, Monaten und Jahren unseres Lebens fort in einer Lebensphase also, in der die für uns wichtigsten Dinge passieren. Es bewertet alles, was Körper und Gehirn tun, entsprechend der Regeln des impliziten (unbewussten) assoziativen Lernens nach "gut/lustvoll/erfolgreich" und damit zu wiederholen bzw. "schlecht/schmerzhaft/erfolglos" und damit zu vermeiden, und legt diese Bewertungen im emotionalen Erfahrungsgedächtnis ab. In dieser Weise formt sich das, was man Charakter oder Persönlichkeit nennt, sehr früh und weitestgehend unbewusst und wird zunehmend resistent gegen spätere Erfahrungen (s. auch unten). Das bewusste Ich sieht sich ab dem vierten Lebensjahr in diese "limbische" Persönlichkeit sozusagen hineingestellt und von ihr getragen.
Unser bewusstes Ich erlebt sich sowohl als Quelle unserer Wünsche, Gedanken, Vorstellungen und Handlungspläne als auch als Verursacher des Handelns, soweit es um so genannte Willkürhandlungen geht, bei denen wir entsprechend das Gefühl haben, wir seien es, die dies gewollt und schließlich auch veranlasst haben. Dies ist der Kern des Gefühls der subjektiven Willensfreiheit.
Auf die Problematik dieses ebenfalls sehr komplexen Begriffs und die Auseinandersetzung um die Existenz oder Nicht-Existenz von Willensfreiheit kann ich hier nicht eingehen, sondern verweise auf das Buch von Henrik Walter (Walter, 1998) sowie auf die letzten Kapitel meines Buches "Fühlen, Denken, Handeln" (Roth, 2001). Aus neurobiologischer Sicht ist Freud in diesem Zusammenhang Recht zu geben: Das unbewusste, limbische Erfahrungsgedächtnis lenkt unser Handeln stärker als unser bewusstes Ich. Grundlage der Beeinflussung des Bewusstseins durch das Unbewusste im Bereich der Gefühle, Wünsche und Vorstellungen ist ein System von Faserbahnen, "ventrale Schleife" genannt, das vom orbitofrontalen und cingulären Cortex zu subcorticalen limbischen Zentren läuft, vor allem zum ventralen Striatum/Nucleus accumbens, von dort aus zum ventralen Pallidum und zur Substantia nigra, und von dort direkt oder über thalamische Umschaltkerne (vor allem den Nucleus mediodorsalis) auf den orbitofrontalen Cortex und den cingulären Cortex zurückwirkt (Abbildung 2).

Abbildung 2: Schematische Darstellung der "ventralen" oder "limbischen" Schleife zwischen corticalen und subcorticalen Zentren der emotionalen Verhaltensbewertung und des Entstehens unserer Wünsche und Absichten. Der limbische Cortex, bestehend aus dem orbitofrontalen und cingulären Cortex, projiziert zum ventralen Striatum (Nucleus accumbens), das seinerseits zum ventralen Striatum projiziert. Beide stehen in enger Verbindung mit der Substantia nigra und dem Nucleus subthalamicus. Auf das ventrale Striatum/Nucleus accumbens sowie das ventrale Pallidum wirken die Amygdala und das Ventrale Tegmentale Areal als Zentren des emotionalen Gedächtnisses, der Hippocampus als Zentrum für das episodische Gedächtnis (Kontextgedächtnis) sowie das basale Vorderhirn (Nuclei septales, diagonales Band von Broca) als Zentrum für Aufmerksamkeitssteuerung ein. Das ventrale Pallidum projiziert zum Nucleus mediodorsalis (MD), der seinerseits zum limbischen Cortex zurück sowie zum ventralen Striatum projiziert. Das ventrale Pallidum und der limbische Cortex haben auch direkte Verbindungen miteinander. Ventrales Pallidum und ventrales Striatum projizieren zum Nucleus tegmentalis pontinoreticularis, von dem der Tractus pedunculo-pontinoreticularis seinen Ausgang nimmt. Weitere Erläuterungen im Text. (Aus Roth, 2001, nach Nieuwenhuys et al., 1991; verändert).
Über diese ventrale Schleife wirken die unbewusst arbeitenden limbischen Zentren auf unser Bewusstsein ein, und zwar in Form des Auftauchens positiver und negativer Gefühle, Gedanken, Assoziationen und Ziele und der Stärke des Wunsches, diese zu verwirklichen. Die genannten subcorticalen limbischen Zentren werden ihrerseits beeinflusst durch den Hippocampus, der zu den genannten Zuständen die kognitiven Details und den Kontext "hinzuliefert", sowie von der Amygala und dem ventralen tegmentalen Areal als den Hauptzentren des emotionalen Erfahrungsgedächtnisses und vom basalen Vorderhirn als dem subcorticalen Zentrum für Aufmerksamkeitssteuerung. Unsere Wünsche werden selbstverständlich auch durch Umweltreize beeinflusst, indem wir bestimmte Dinge oder Vorgänge sehen, hören, riechen usw., aber diese müssen, um echte Handlungsantriebe zu werden, auf eine interne Motivationslage treffen. Viele Umweltgeschehnisse lassen den einen "völlig kalt", während sie auf den anderen höchst stimulierend wirken, und dieser Sachverhalt wird durch das individuelle emotionale Erfahrungsgedächtnis bestimmt.
Charakteristisch ist, dass die corticale Bewusstseinsebene die starke Beeinflussung durch die subcorticalen limbischen Ebenen nicht wahrnimmt oder sie gar leugnet. Entsprechend scheinen unsere Gedanken, Absichten, Wünsche und Handlungspläne von nirgendwo her "aufzutauchen", sie kommen uns "plötzlich", oder wir schreiben sie uns selbst, d.h. dem Ich, zu, dessen Status gleichzeitig ihm/uns rätselhaft bleibt. Die Erklärung für diesen Vorgang besteht darin, dass das bewusste Ich die Herkunft dieser intentionalen Empfindungen nicht zu den subcorticalen limbischen Zentren zurückverfolgen kann. Ganz offenbar ist es dem bewussten Ich auch unerträglich, die sich selbst als dem Verursacher zugeschriebenen Handlungsweisen als "unerklärt" stehen zu lassen. Das Ich konfabuliert, d.h. es liefert aus Sicht des Beobachters Pseudo-Erklärungen, und zwar in der Regel solche, die dem Selbstwertgefühl und den Erwartungen der sozialen Umgebung am besten entsprechen. Die dabei unvermeidlich auftretenden Diskrepanzen zwischen Tun und verbaler Erklärung werden durch ständige Abänderungen der verbalen Erklärungen vorübergehend behoben.
Diese konfabulatorischen "Rettungsmaßnahmen" sind relativ unproblematisch, so lange sie nicht die innere Konsistenz und Kohärenz des unbewussten verhaltenssteuernden Apparates nachhaltig stören oder schwere Konflikte mit der sozialen Umgebung hervorrufen. Tritt jedoch hiervon das eine oder andere auf und wird die Erklärungskapazitäten des bewussten Ich überschritten, dann kommt es entweder zum bewussten "Abstandnehmen" des Ich von den Handlungen, wie es bei Neurosen und Zwangshandlungen der Fall ist, oder zu krassen Realitätsverbiegungen, die sich in Psychosen äußern. In jedem Fall ist dann die oben genannte "ventrale Schleife" in ihrer Funktion gestört: Der neurotische Patient erlebt die ihn beherrschenden Wünsche nicht mehr als die eigenen, sondern als "Zwang", und der psychotische Patient interpretiert die Geschehnisse in seiner Umwelt und seine eigenen Handlungen durch die "Zwangsbrille" seines gestörten Unbewussten.
Das Dogma der heutigen Neurowissenschaften lautet, dass alle Leistungen des Gehirns seien sie perzeptiver, kognitiver, affektiv-emotionaler, exekutiver oder motorischer Art Funktionen von Neuronen-Netzwerken sind. Die hierbei entscheidenden Faktoren sind zum einen die morphologisch-physiologischen Eigenschaften der Neurone als Knotenpunkte der Erregungsverarbeitung, zum anderen die neurophysiologischen und neurochemischen Eigenschaften der Synapsen, von denen es im menschlichen Gehirn ca. 1 Trillion (1015) gibt. Psychische Erkrankungen beruhen aus dieser Sicht auf dysfunktionalen Veränderungen von Neuronen-Netzwerken insbesondere im limbischen System, wobei diese Veränderungen auf genetische Defekte, Erkrankungen oder auf vorgeburtlich, frühkindlich oder in späterem Lebensalter erlittene Schädigungen zurück gehen können. Diese Veränderungen drücken sich zum Beispiel in einer abnormen Erhöhung oder Erniedrigung von Neuromodulatoren (z.B. Dopamin oder Serotonin) oder Neuropeptiden (z.B. Arginin-Vasopressin, Oxytocin) in bestimmten limbischen Zentren aus. Dass psychische Erkrankungen auf derartige Veränderungen in Neuronen-Netzwerken zurückzuführen ist, sagt freilich nichts über die Ursachen aus, denn Veränderungen in der Verarbeitungseigenschaften von Neuronen-Netzwerken bedeuten erst einmal nichts anderes als Kommunikationsstörungen zwischen Gehirnzentren.
Untersuchungen mit bildgebenden Verfahren (PET, fMRI) zeigen in der Tat, dass bei psychisch Kranken im Vergleich zu psychisch Gesunden limbische Zentren in ihrem Aktivitätszustand deutlich erhöht oder erniedrigt sind. Ziel jeder Psychotherapie muss es entsprechend sein, die Psyche des Patienten dadurch zu verändern, dass die Fehlfunktionen subcorticaler limbischer Netzwerke behoben werden. Dies ist jedoch allein schon aus neurobiologischer Sicht mit erheblichen Schwierigkeiten verbunden.
Man muss nämlich davon ausgehen, dass corticale und subcortical-limbische Netzwerke sich grundlegend in der Veränderbarkeit ihrer synaptischen Kontakte unterscheiden. Corticale Netzwerke sind schnell veränderbar, und zwar etwa im Sekundentakt, der auch der Takt des Bewusstseins ist. Diese Netzwerke können also in "Sekundenschnelle" Informationen aufnehmen, sie mit vorhandenen Informationen aus den verschiedensten funktionsspezifischen Gedächtnissen verbinden und so bereits bestehende Netzwerke verändern oder neue Netzwerke schaffen. Schnelligkeit, hohe Verarbeitungskapazität und hohe Verknüpfungsplastizität zeichnen also das bewusstseinsfähige cortico-hippocampale System aus. So können wir den Inhalt eines Satzes sofort verstehen und einen Befehl sofort befolgen ("bleib stehen!"), sofern unser limbisches System dagegen keinen Einspruch erhoben hat.
Allerdings ist bisher nicht genau bekannt, aufgrund welcher Eigenschaften und Mechanismen corticale Synapsen diese schnellen Veränderungsmöglichkeiten besitzen. Sehr wahrscheinlich geht es dabei um Veränderung der Leitfähigkeiten spannungsabhängiger oder ligandgebundener chemischer Synapsen oder den Einsatz elektrischer Synapsen ("gap junctions"). Anatomische Veränderungen der synaptischen Verknüpfungsstrukturen, z.B. über die Bildung so genannter Dornensynapsen, sind in diesem Kontext unwahrscheinlich; dies geschieht wohl vor allem bei der Bildung des Langzeitgedächtnisses (für Einzelheiten siehe Roth, 2001).
Subcorticale limbische Netzwerke hingegen haben eine nur beschränkte Informationsverarbeitungskapazität, d.h. sie sind nicht in der Lage, große Mengen von Detailinformationen schnell zu verarbeiten und sinnvoll miteinander zu verknüpfen. Objekte und ihre Kontexte werden nicht in ihren Einzelheiten, sondern anhand charakteristischer Merkmale erkannt; insbesondere ist der intermodale Informationstransfer stark eingeschränkt. Insgesamt gehen die neuronalen Umverknüpfungen in limbischen Zentren deutlich langsamer vor sich, was darauf hindeutet, dass hier vor allem so genannte metabotrope Rezeptoren eine Rolle spielen. Diese wirken steigernd oder hemmend über intrazelluläre Signalketten längerfristig auf die Übertragungseigenschaften von Synapsen ein, indem sie über Genexpression zum Beispiel die Vermehrung oder Verminderung von Rezeptoren oder die Neubildung oder das Einschmelzen von Dornensynapsen bewirken. Entsprechend vollziehen sich emotionale Lernvorgänge und Gedächtnisbildungen vergleichsweise langsam, im charakteristischen Gegensatz zu schnellen cortical-kognitiven Informationsverarbeitungsprozessen. Besonders langsam vollziehen sich spätere Umlernprozesse. Wenn wir erst einmal mehrfach erfahren haben, dass irgendetwas schlecht für uns ist, dann dauert es meist sehr lange, bis sich durch nachfolgende positive Erlebnisse die negative Erfahrung in ihr Gegenteil verkehrt. Die ersten Erlebnisse sind offensichtlich emotional stets besonders wirksam.
Wir müssen gleichzeitig davon ausgehen, dass es während der Individualentwicklung bei limbischen Lern- und Gedächtnisbildungsprozessen "sensible" Phasen gibt, wie sie aus der Verhaltensbiologie bei Prägungsprozessen bekannt sind. Hierbei werden Netzwerkstrukturen und -funktionen so verändert, dass sie gegen spätere Veränderungen relativ resistent sind. Solche "sensiblen" Phasen scheinen während der Gehirnentwicklung im Mutterleib, kurz nach der Geburt und in den ersten Lebensjahren stattzufinden. Dies würde bedeuten, dass Veränderungen außerhalb dieser sensiblen Phasen mit zunehmendem Alter immer schwerer vonstatten gehen. Dies würde insbesondere für limbische Netzwerke gelten, die aufgrund traumatischer Ereignisse, d.h. aufgrund starker affektiver und emotionaler Belastungen in ihren Strukturen festgelegt wurden. Entsprechend hoch müsste die emotionale Aktivierung (z.B. Lebenskrisen oder eine fordernde neue Partnerschaft) sein, um diese Prägungen wieder aufzuheben, damit sich an unserer Persönlichkeit überhaupt noch etwas ändert.
Entsprechend bleiben bloße Appelle an die Einsicht wirkungslos, denn sie aktivieren allein die Netzwerke des bewusstseinsfähigen cortico-hippocampalen Systems; dieses hat aber auf die verhaltensrelevanten limbischen Netzwerke keinen wesentlichen oder einen nur indirekten Einfluss. Eine Veränderung des cortico-hippocampalen Systems verändert unser deklaratives Gedächtnis, nicht aber unser Verhalten. Dies bedeutet generell, dass Einsicht allein nicht zu einer Verhaltenänderung führt, und dass man sich über Einsicht nicht selbst therapieren kann.
Der Psychotherapeut hingegen kann mit geeigneten therapeutischen Mitteln, insbesondere mithilfe der Erzeugung eines "emotionalen Aufruhrs", auf das Unbewusste des Patienten einwirken und damit Veränderungen subcorticaler limbischer Zentren bewirken. Diese therapeutischen Einwirkungen lösen die gesteigerte Ausschüttung bestimmter Neuromodulatoren und Neuropeptide aus. Wie dies in einer erfolgreichen Psychotherapie im Einzelnen geschieht, ist allerdings noch unbekannt.
Manche Emotions-Neurobiologen, z.B. Joseph LeDoux, bezweifeln, dass das limbische Erfahrungsgedächtnis überhaupt vergisst oder umlernen kann, wenn es erst einmal emotional geprägt wurde. LeDoux (1998) nimmt an, dass im Laufe einer Therapie das bewusstseinsfähige cortico-hippocampale System nicht in das limbische emotionale Erfahrungsgedächtnis eingreift (was es eventuell gar nicht kann), sondern an diesem Erfahrungsgedächtnis vorbei seinen eigenen Zugang in die Handlungssteuerung gewinnt. Dies würde bedeuten, dass man sich zum Beispiel in Angst- und Furchtsituationen zunehmend beruhigt, ohne dass die tiefen Quellen der Angst und Furcht beseitigt sind; sie sind lediglich "übertüncht". Tritt nach einer Serie positiver Erfahrungen wieder das frühere negative Ereignis auf, dann ist es so, als sei die frühere Angst nie wirklich geschwunden.
Zweifellos gibt es derartige Fälle, in denen durch Therapiemaßnahmen die Quellen emotional-affektiver Störungen lediglich übertüncht, aber nicht beseitigt sind. In anderen Fällen kommt es indes doch zu einer langfristigen Veränderung in limbischen Netzwerken, so dass die Patienten sich der gefährlichen Situation aussetzen können, ohne einen Rückfall zu erleben. Diese Fragstellung wird Gegenstand intensiver neurobiologischer Untersuchungen sein.
Zwischen den Neurowissenschaften und der Psychoanalyse hat Jahrzehntelang eine erbitterte Feindschaft bestanden. Für die Psychoanalytiker war die Hirnforschung das abschreckende Beispiel reduktionistischen Vorgehens oder (wie Freud zuweilen meinte) eine völlig überflüssige empirische Beigabe zur "wahren Lehre". Für die Neurowissenschaftler (ebenso wie für viele andere empirisch-experimentell arbeitenden Wissenschaftler) galt die Psychoanalyse als Inbegriff unwissenschaftlicher Theoriebildung des Psychischen, während man gleichzeitig selbst überhaupt nicht wusste, wie man mit empirisch-experimentellen Mitteln das Psychische angehen könnte. Dies hat sich in den letzten Jahren geändert, und die Strukturen und Funktionen des limbischen Systems bzw. die neuronalen Grundlagen der Affekte und Emotionen sind zu einem hochaktuellen neurowissenschaftlichen Forschungsgegenstand geworden. Zumindest kann die Neurobiologie heute in groben Zügen angeben, "wie das Gehirn die Seele macht".
Im Laufe dieser Forschung haben sich Resultate und Einsichten ergeben, die wie ich in diesem Aufsatz umrisshaft darzustellen versuchte die Lehre Freuds in einigen wichtigen Punkten zu bestätigen scheinen. Diese lauten: (1) Das Unbewusste bestimmt weitgehend das Bewusstsein; (2) das Unbewusste entsteht ontogenetisch vor dem Bewusstsein; es legt sehr früh die Grundstrukturen der Weise fest, wie wir mit uns und unserer Umwelt umgehen; (3) das bewusste Ich hat keine oder eine nur geringe Einsicht in die unbewussten Determinanten des Erlebens und Handelns; (4) das emotionalen Erfahrungsgedächtnis hat das erste und das letzte Wort, nämlich beim Entstehen unserer Wünsche und Handlungsabsichten und bei der Letztentscheidung über die Realisierung dieser Wünsche und Absichten.
Es ist der zukünftigen Forschung vorbehalten, diese sich abzeichnende Konvergenz Freudscher psychoanalytischer Grundanschauungen und neurowissenschaftlicher Erkenntnisse zu bekräftigen oder abzuschwächen. Damit wäre auch ein wichtiges Wort über die wissenschaftliche Absicherung der psychoanalytischen Therapie gesprochen. Es muss deshalb von großem Interesse für die Psychotherapie und die Psychoanalyse sein, einer Kooperation mit den Neurowissenschaften aufgeschlossen gegenüberzustehen.
Aggleton, J.P. und M.W. Brown (1999): Episodic memory, amnesia, and the hippocampal-anterior thalamic axis. Behavioral Brain Sciences 22: 425-489 (Zit.)
Anderson, S.W., A. Bechara, H. Damasio, D. Tranel und A.R. Damasio (1999): Impairment of social and moral behavior related to early damage in human prefrontal cortex. Nature Neuroscience 2: 1032-1037 (Zit.)
Bechara, A., D. Tranel, H. Damasio, R. Adolphs, C. Rockland und A.R. Damasio (1995): Double dissociation of conditioning and declarative knowledge relative to the amygdala and hippocampus in humans. Science 269: 1115-1118 (Zit.)
Bechara, A., Damasio, H., Tranel, D. and Damasio A.R. (1997): Deciding advantageously before knowing the advantageous strategy. Science 275: 1293-1295 (Zit.)
Damasio, A.R. (1994): Descartes' Irrtum. Fühlen, Denken und das menschliche Gehirn. List, München (Zit.)
Davidson, R. J. und W. Irwin (1999): The functional neuroanatomy of emotion and affective style. Trends in Cognitive Sciences 3: 11-21 (Zit.)
Goschke, T. (1996): Gedächtnis und Emotion: Affektive Bedingungen des Einprägens, Erinnerns und Vergessens. In: D. Albert und K.-H. Stapf (Hrsg.), Enzyklopädie der Psychologie. Themenbereich C, Serie II, Bd. 4 Gedächtnis. Hogrefe Verlag für Psychologie, Göttingen, Bern, Toronto, Seattle, S. 603-692 (Zit.)
Goschke, T. (1997): Implicit learning and unconscious knowledge: Mental representation, computational mechanisms, and neural structures. In K. Lamberts und D. Shanks (Hrsg.), Knowledge, concept and categories. Hove, UK, Psychology Press, S. 247-333 (Zit.)
LeDoux, J. (1998): Das Netz der Gefühle. Wie Emotionen entstehen. Carl Hauser Verlag München-Wien (Zit.)
Lurija, A.R. (1991): Der Mann, dessen Welt in Scherben ging. Rowohlt, Reinbek (Zit.)
Markowitsch, H.J. (1999): Gedächtnisstörungen. Kohlhammer, Stuttgart (Zit.)
Newen, A. und K. Vogeley (2000): Selbst und Gehirn. Mentis, Paderborn (Zit.)
Panksepp, J. (1998): Affective Neuroscience. The Foundations of Human and Animal Emotions. Oxford University Press, New York, Oxford. (Zit.)
Petrides, M. und D. N. Pandya (1999): Dorsolateral prefrontal cortex: comparative cytoarchitectonic analysis in the human and the macaque brain and corticocortical connection patterns. Europ. J. Neuroscience 11: 1011-1036 (Zit.)
Rolls, E.T. (1999): The Brain and Emotion. Oxford University Press, New York, Oxford. (Zit.)
Roth, G. (1996): Das Gehirn und seine Wirklichkeit. 2. veränderte Aufl. Suhrkamp, Frankfurt (Zit.)
Roth, G. (2001): Fühlen, Denken, Handeln. Wie das Gehirn unser Verhalten steuert. Suhrkamp, Frankfurt (Zit.)
Sacks, O. (1987): Der Mann, der seine Frau mit einem Hut verwechselte. Rowohlt, Reinbek (Zit.)
Spanagel, R. und F. Weiss (1999): The dopamine hypothesis of reward: past and current status. Trends in Neurosciences 22: 521-527 (Zit.)
Tulving, E. und H. J. Markowitsch (1998): Episodic and declarative memory: Role of the hippocampus. Hippocampus 8: 198-204 (Zit.)
Walter, H. (1998): Neurophilosophie der Willensfreiheit. Mentis, Paderborn (Zit.)