Neurology, emotion organs
Introduction
What in this article is denoted as the "emotion organs" are in principle all structures lying between cortex (the curls on top in the illustration on the right) and the brainstem (the lower yellow area). The boundary with the cortex is well-definable: this is the corpus callosum, the large area with connections between the right hand and left hand half of the cortex (the upper yellow area). That with the brainstem is somewhat less clear, but a suitable choice would be the point where the brainstem splits into a left hand and right hand branch.The point with the present choice is that it does not correspond to the more usual ones. Of which there are several so making a choice is necessary anyway. A choice that is also hampered by the continuously enhancing knowledge of the working of the brain.
Because of these difficulties, this article starts with a repeat of a part of the end of the description of the brainstem
 (in Dutch), so that it can be read independently.
As with this precursor, the primary intention is to show the goal of all these
structures as far the personal "user" is concerned, and provide as much technical
detail as necessary but not more.
(in Dutch), so that it can be read independently.
As with this precursor, the primary intention is to show the goal of all these
structures as far the personal "user" is concerned, and provide as much technical
detail as necessary but not more.The neurological naming system comes from early anatomy and is based on appearance and location, and has little to no relation to actual functionality. And is quite inconsistent and varies considerably. Another goal of this article is to give enough information in order to be better able to read through this in other literature.
Brainstem
The brainstem builds upon the work already done in the spinal chord. The latter directly controls the limbs, and together the two control all bodily functions that are executed unconsciously.This is a complicated task and there must be some organization in it, and nature does this preferably step by step: a specific neurological structure takes care of one specific task. If two tasks have to be coordinated, this task of coordination is done by another specific structure. If more than two tasks have to be coordinated, then, as a rule, even this is done one step at a time.
The cognitive human mind knows that this is due to the fact that three-tiered systems tend to be less stable - with two systems it is much easier to achieve an equilibrium. The evolutionary reason is that any new functionality must be build while the present structures go on working - a pause during building work is not an option in real live.
This process of "building upon" leads to a string of structures in the brainstem, see the illustration below (from here
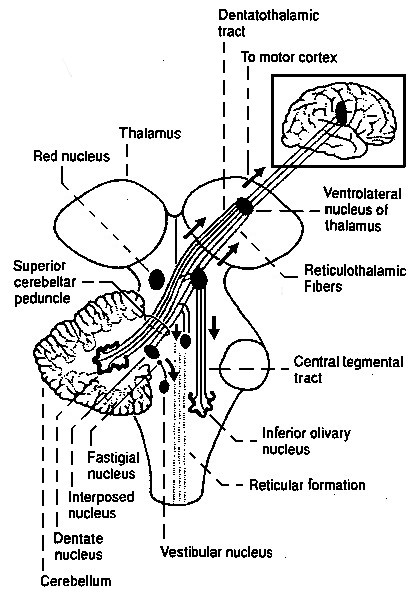 |
This illustration should be "read"to be "read" from the bottom upwards (!). So the first larger structure is the olivary nucleus, followed by others like the cerebellum and the red nucleus, with many smaller ones in between,
The olivary nucleus is also the first one having a clearly laminated structure, of the type generally called a "neural network", having the folded look familiar from the cortex. The olivary nucleus probably does the first larger-scale coordinating work.
It is followed by the much larger cerebellum, which is probably an averaging-device: a mechanism that learns from lots of previous behaviour the fine-control of behaviour
The last nucleus undisputedly within the brainstem is the red nucleus, that is generally associated with "gait": the coordination of multiple limbs for the propagation of the organism.
The basis of all this control is the neuron: a cell specialized in the propagation of signals and looking like a wire-with-switch:
 |
In its primitive form, in the famous (simplified) example, it is at one end connected to sense organs in the knee and at the other end to muscles in the leg. Tap the knee, and the leg stretches automatically.
In the spinal cord there are also lots of ganglia, conglomerates of neurons, that coordinate planned movement with actual movement, that coordinate the movement of separate parts of a limb to that of the limb as a whole, etcetera.
To do all this, the neurons have to communicate with each other. This is done through biochemical substances called neurotransmitters. The one involved in the basic knee reflex is glutamate, the neurotransmitter that makes the neuron "fire" - and excite a muscle, or a neighbouring neuron.
For the process of coordination there is also a neurotransmitter that blocks, or inhibits, neighboring neurons, called GABA. Living nature has a strong preference for developing systems with "forces" and "counter forces", probably because they lead to more stable yet flexible forms of equilibrium.
The state of equilibrium has many advantages, allowing the optimization of the involved sub processes to the normal circumstances. However, nature serves all kinds of surprises, many of the unpleasant type that a live form should avoid. And get away from this situation as quickly as possible, by humans called an "emergency". The emergency response must be extra fast and powerful. But this should be limited to the emergency situation, or else it would take so much energy and supplies that the regular food intake is not sufficient.
This is one of the things added to the neurological system by the middle part of the brainstem: the production of a neurotransmitter that excites the system for emergencies, norepinephrine (or noradrenalin), and one that puts the system back to equilibrium values: serotonin. That of norepinephrine is called the locus coeruleus and is located in middle part of the brainstem, and for serotonin there are seven of these ("raphe") "nuclei", with separate ones serving the spinal cord, brainstem, and upwards lying structures.
Where as glutamate and GABA are an intrinsic part of the neuron-chemistry, norepinephrine and serotonin are synthesized in neurons dedicated to this purpose, grouped at the named locations. These neurotransmitters work on the connecting points of the other neurons, the so-called "synapses".
All this is the executive part of what is described as the "fight or flee" situations.
Another possiblity to preserve energy consumption i.e. demand for food, is to apply the emergency state only when strictly needed. But in nature these kind of circumstances will vary during life. So the thing to do is to apply this state depending on the kind of environment. This is what humans call "learning". This means that certain behaviour must be stimulated, and other behaviour discouraged.
This is executed by two more neurotransmitters, this time in the top of the brainstem. One quite famous: dopamin, and one usually ignored in this kind of descriptions: acetylcholin. Dopamin is well-known because it makes you do things. And rather urgently, at that. All or almost all addictive behaviour is related to dopamine.
By the token of "force - counterforce", acetylcholin would be the stimulus to avoid behaiour, which explains why it is rarely noticed: its results are not directly observable.
Here is an overview of the relevant parts of the brainstem where these four modulating neurotransmitters are produced - the green area is called tegmentum and the yellow-shaded part in its front is the ventral tegmental area, or VTA (original illustration from the site of Ben Best
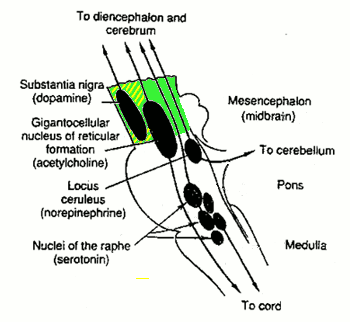 |
Usually substantia nigra ("black substance") and locus coeruleus ("blue area") are grouped into the so-called "limbic system", because they produce the substances that are used by the limbic system. This seems to make little sense, because then one should also include the other two. A reason for the alternative division made here.
Another departure from normality in divisions is the inclusion of the pituitary and pineapple glands with the brainstem. These produce the neurotransmitters that get into the bloodstream, then called "hormones". Here the argument is used the other way around: because the glands produce the equivalent of neurotransmitters for the bloodstream, and the production of the basic neurotransmitters takes place in de brainstem, the glands are also considered to be part of the brainstem. Which argument then also applies to the hypothalamus, that controls the pituitary gland.
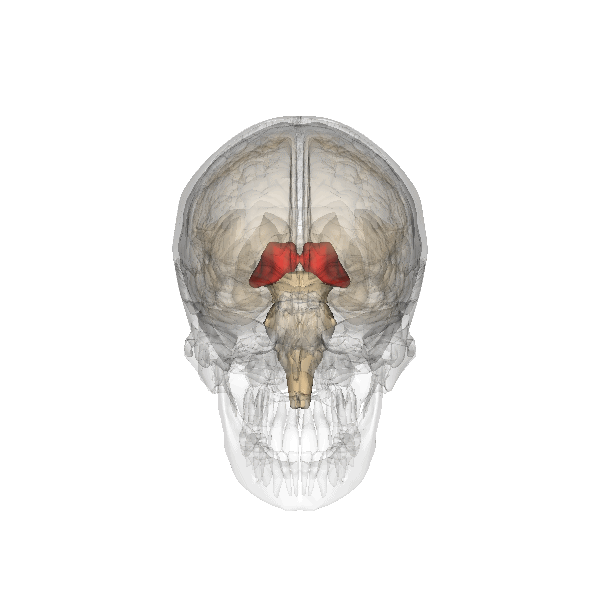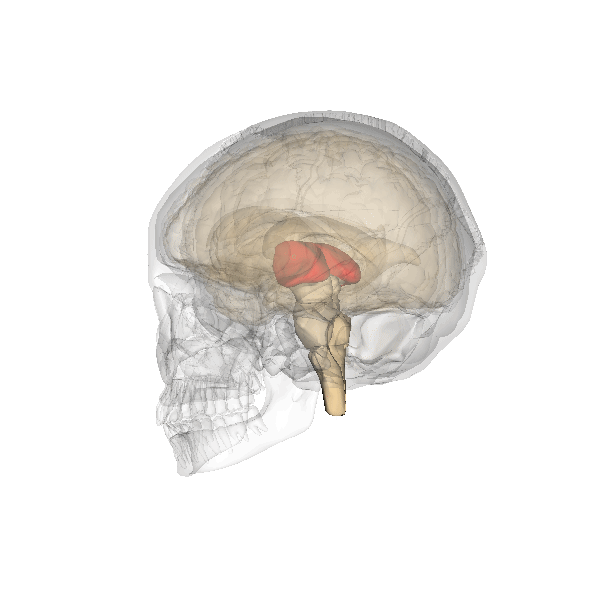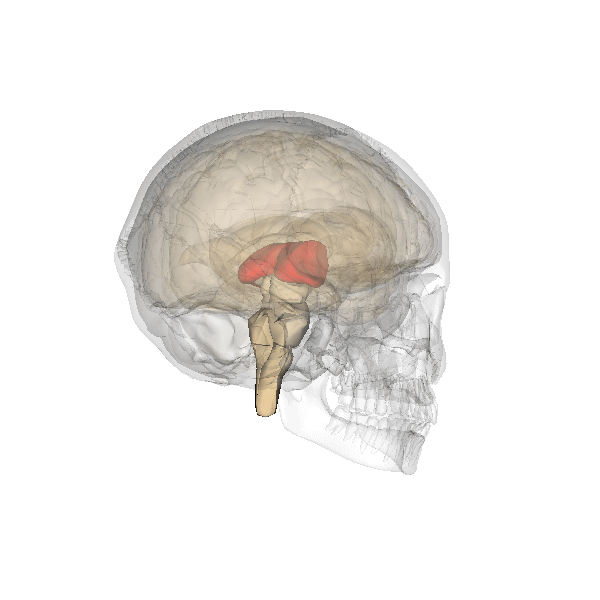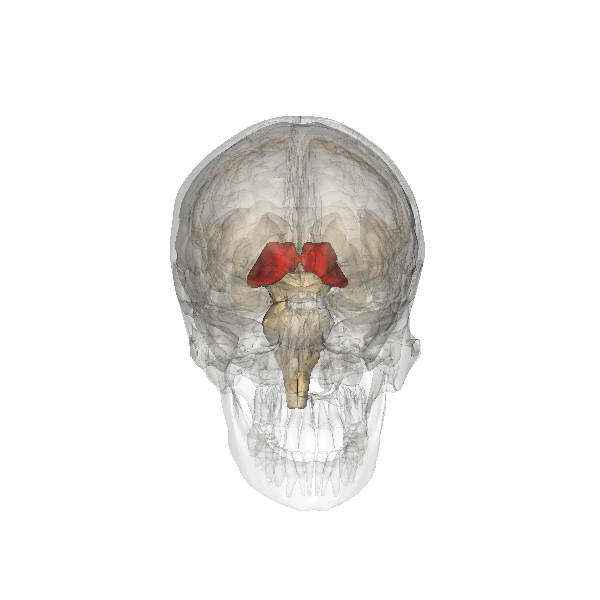
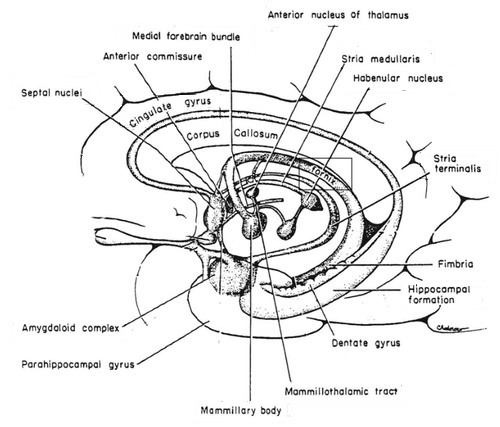
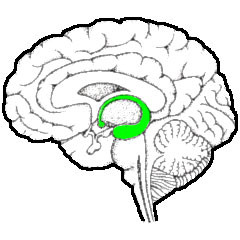
For which there are physiological arguments too. For example: all emotion organs come in a left sided and a right sided version, the brainstem splitting visibly at the top. For both hypothalamus, pituitary and pineapple gland there is only one, lying in the middle, between the two structures called thalamus, that are more or less the functional border between brainstem and emotion organs, see the following illustration:
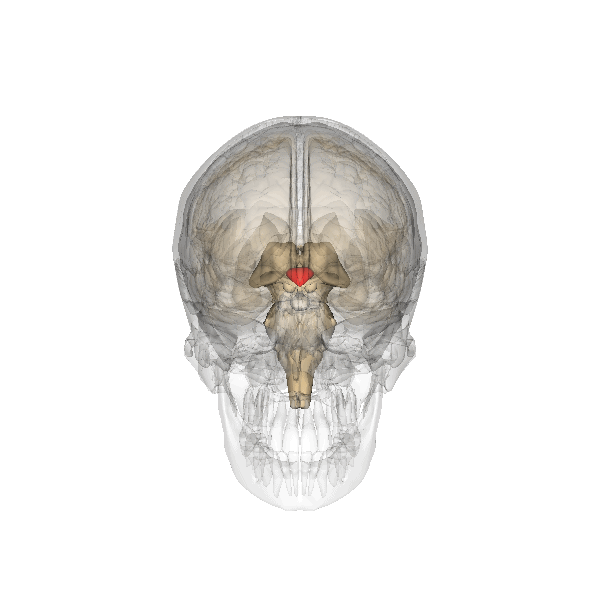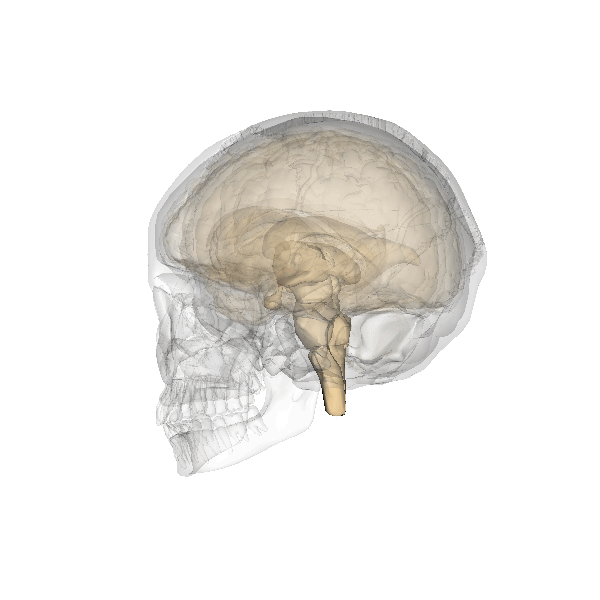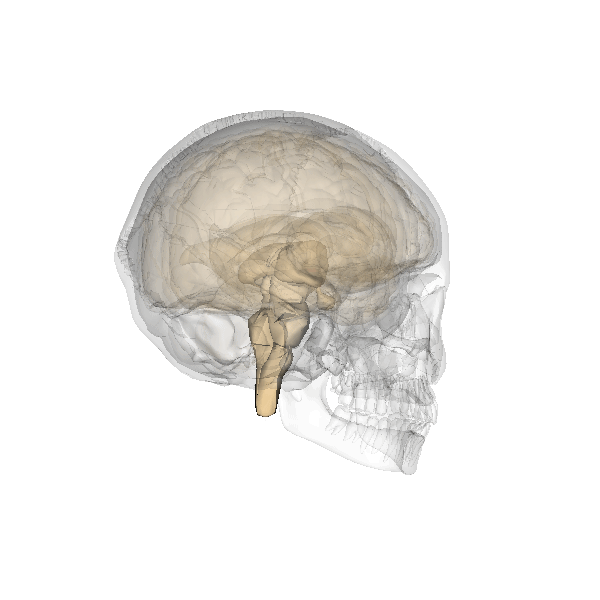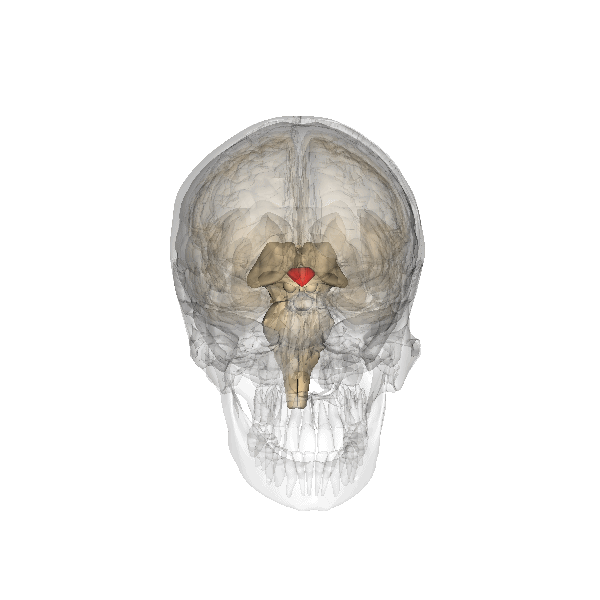 |
The hypothalamus is the red-colored element, and the pituitary gland (English: pituitary gland or pituitary) is the gray "ball" which it is attached to it.
So where for example the neurological system is put into the state of alarm by the release of norepinephrine or noradrenalin, for the rest of the body this is done by the release of adrenelin by the pituitary gland into the bloodstream. And the same goes for other neurotransmitters and the associated processes.
In this context, the brainstem also increasingly has to do things that humans call "decisions". Those associated with emergencies are known as those of "fight or flee", but in fact the choice is wider: "fight, flight, freeze or go for it" - as signified by there being four modulating neurotransmitters that can be associated with these choices.
In the brainstem there must be structures that make preliminary decisions on this, as exemplified by the phenomenon of "fainting": when the stress on the system gets to certain levels, the whole system is shut down. And there is little doubt that this is done in the brainstem, since is also involves the shutdown of all muscles etcetera. By what structures this is done, is not known (it might be the peri-aquaductal grey
As the fainting example shows, these decisions are usually quite "digital" in nature: it is "on" or "off". So there is room for improvement. For which nature developed higher structures. The first of which beyond the brainstem is the thalamus, which at present has a bridging function, passing information from the brainstem to the higher structures and the other way around. The central position of the thalamus is illustrated below:
|
But this bridging function may be a later role, where the thalamus initially was the point where the first more subtle decisions based on more observational information were made.
The connections to and from the thalamus are shown in the illustration below (from here
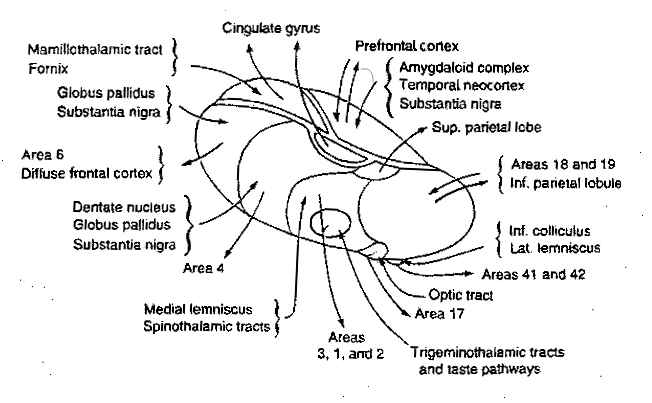
|
Some of its structures could not be readily included in this illustration, among which are two important ones: the body of the thalamus is cloaked for about two-thirds by a shell-like nucleus called the reticular nucleus that is able to activate or deactivate the nuclei below it. And in its interior, that is largely filled with internal connections, there lie two nuclei along the length of the body, see the next illustration:
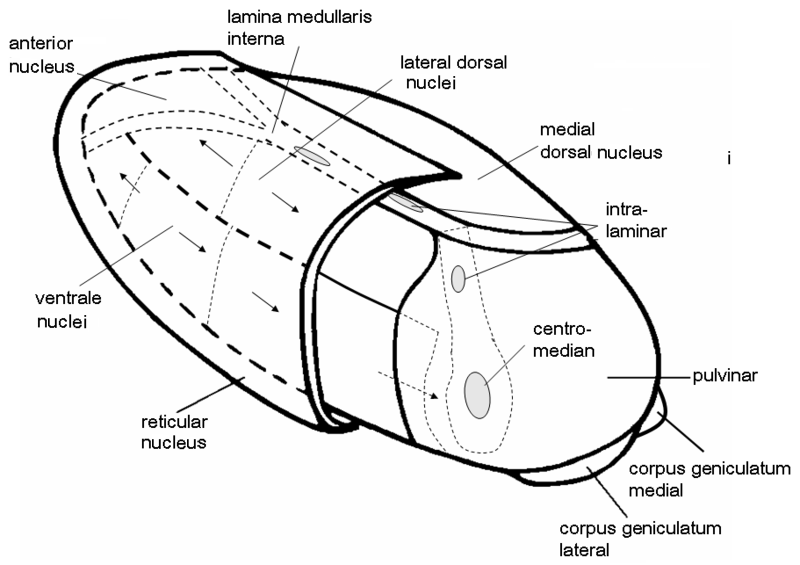 |
The larger one, the centromedian nucleus, is connected to the next structural layer: the basal ganglia. So probably the centromedian nucleus also has a centralizing function.
The existence of this cloaking reticular nucleus and that of a central nucleus which is sending signals to the emotion organs supports the assumption that the thalamus has an integrating function with respect to the miscellaneous aspects of the perceived reality together with the already existing physical and neurological reactions to it.
The thalamus thus probably plays a role in creating a complete picture of the reality from its various individual aspects. This is a continuation of earlier forms of integration and coordination that take place within the brainstem.
Presently quite well-known is the phenomenon of selective attention as illustrated by a group of people playing a ball game, into which a person dressed as a gorilla walks in, see the picture below:
 |
More than half of the participants do not notice the gorilla, busy as they are playing the game.
This process of selective attention (and there are numerous other examples from daily life) must take place at quite a fundamental level, in order for the basic processes of observation i.e. the signals of the sensory organs, to be influenced. It is quite likely that this process of selective attention takes place in and around the thalamus, the reticular nucleus being the one that does the disabling of the parts of the observational process.
All this supports the assumption that in the thalamus the first total integration of observations and body responses to those observations takes place - the first "virtual reality". It is this information that can be used to assess the effects of behaviour. With this information you can build a system to decide on future behaviour: "Do we do this, or we do that?" So making decisions based on this information, but, compared to what the brainstem does, already much more subtle ones.
To which more subtle decision-making, the rest of the brain makes even more contributions, starting with the next structural layer: emotion organs.
Emotion organs, globally
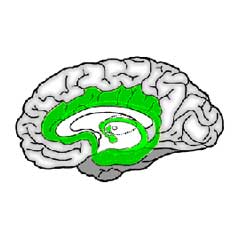
For the structures lying directly beyond the brainstem there are several designations: viscereal system, limbic system, basal ganglia, basal forebrain, which partially overlap. And sometimes they include structures from the brainstem, notably those that produce neurotransmitters. Here, as stated in the introduction, we cut through these woods, by taking all structures beyond the brainstem until arriving at the cortex as the "emotion organs", see the illustration below:
 |
Within the green-colored area the substructures are separated by clearly drawn lines, but that is done for clarity: in reality the emotion organs form a compact unit with sometimes quite vague transitions. Former classifications drew purely on anatomical characteristics (color and the like), but nowadays it is more and more the type of neurons in the structures that determine its classification.
There is little argument in the standard literature on which is the first one of these subgroups: the basal ganglia. The same term is going to be used here, but with an extended population.
Basal ganglia, part one
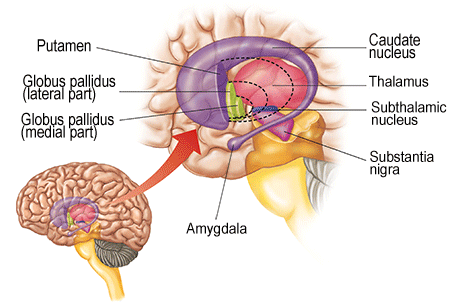
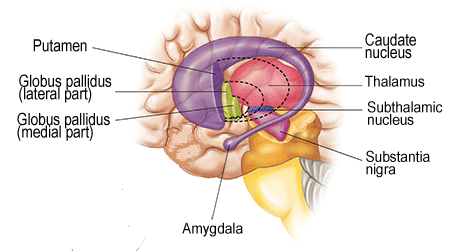
The basal ganglia are the first structures one encounters starting from the thalamus going upwards and sideways. As stated above, the thalamus seems to be the location of the first more or less complete view of the world - the virtual reality of the brain.The following structures then can use this view to make better judgements on what to do than the brainstem does, working on this information through a sequence of structures. The thalamus connects to the first of these, the global pallidus, at its inner layer. This connects to an outer layer of the globus pallidus, this to the putamen, and this to the caudate nucleus, all tightly packed together in a complicated whole that is difficult to depict, see the illustrations below (first from here
 , second from here
, second from here
 , third from here
, third from here
 ):
):
 |
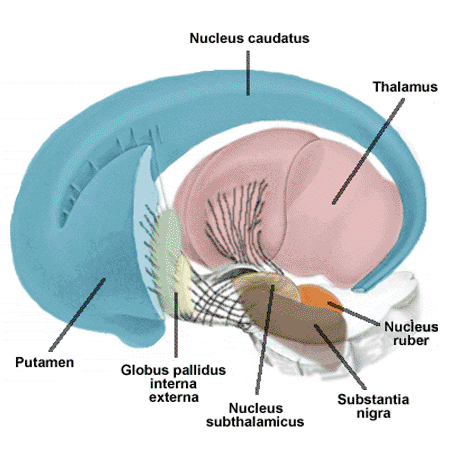 |
These are sideways views with the nose to the left. In these types of illustrations the connections are usually omitted (directly above they are visible schematically) - e.g. the spaces between the various components are filled completely with axon bundles going to and fro other structures.
The third image is a vertical cross-section through the separation of left and right hemispheres, seen from the front:
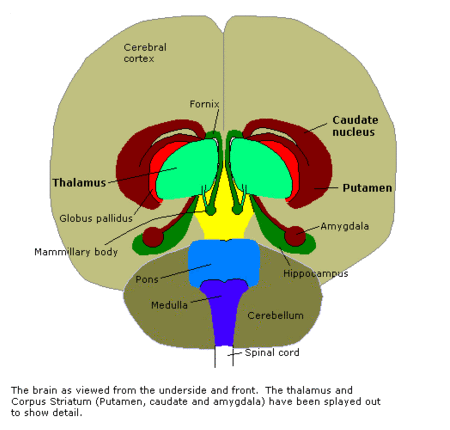 |
This clearly shows the "shell-like structure": first, the thalamus, surrounded by the globus pallidus ("pale sphere"), then follows the putamen ("peach-like nucleus") and wrapped around this whole is draped the tail of the caudate nucleus ("nucleus with tail") - its head is attached at the front of the putamen, see the 3D-illustration below:
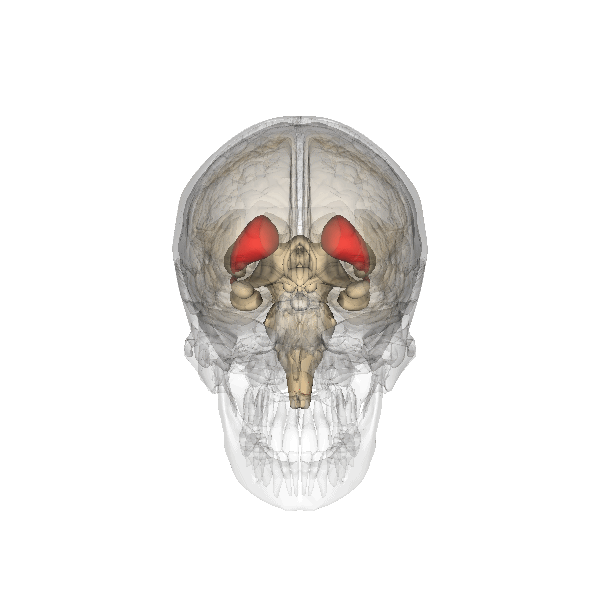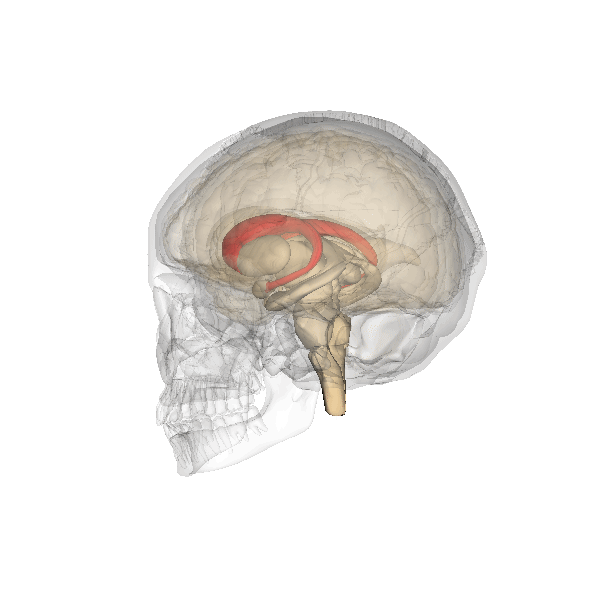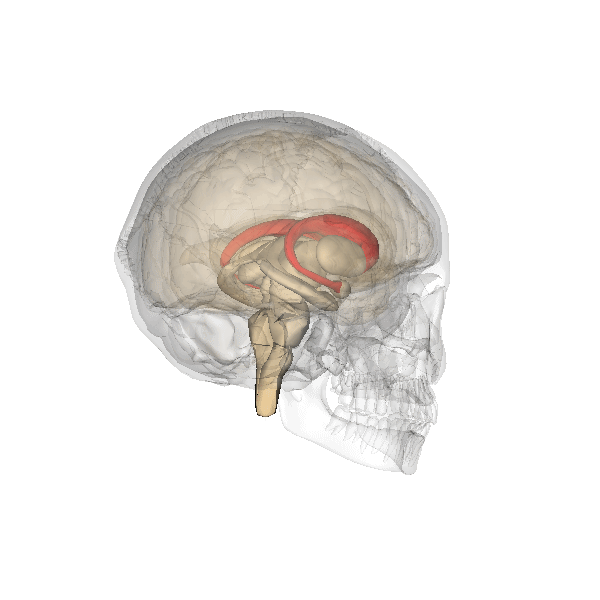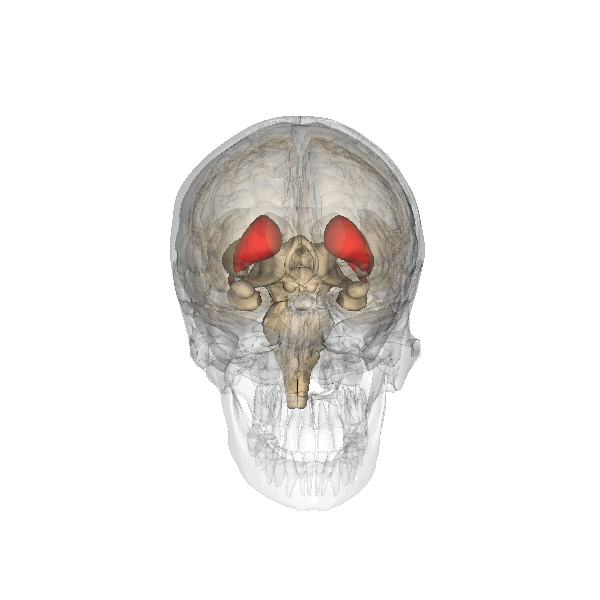 |
The combination of putamen and caudate nucleus is also referred to as the striatum, according to the more or less striped appearance that they share - or more specifically: the dorsal striatum, dorsal meaning in (standing) humans: on the backside. In this convention the global pallidus is called the dorsal pallidum.
Due to there combined shape in cross section, the combination of globus pallidus plus putamen is also referred to as lentiform nucleus, see the illustration below (detail of Gray 742 - this and subsequent etching of anatomy atlas of Gray
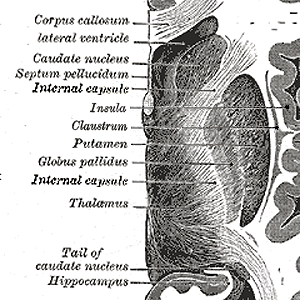 |
Clearly visible is the character of it being a sequence, starting with the thalamus, its connections denoted as internal capsule (other connections run through this area too), globus pallidus (its two layers just visible), putamen, and then, anatomically lying outside the emotion organs: claustrum and insula, the last one being a part of the cortex. It is difficult to look at this combination without imagining a functional relation between them,on which some remarks later.
The previous largely follows standard descriptions.
As important as determining the different structures and substructures, are the connections between them. Here there is an important and largely disregarded problem: the neurons have two kinds of connections: firstly the axons, a dedicated output used for longer distance connections. The axons have an insulation-cover that makes them look lighter, and they come usually in bundles, making them clearly visible.
The other kind is that of neurons with no clearly visible axon and that connects neighbour to neighbour. This kind of neuron is called an interneuron, and they are in the majority in the brain - they do the "calculating". Their connections do not readily appear to the human investigator. The globus pallidus interna, externa, putamen en globus pallidus are clearly connected in this way. And clearly in a stream of information and control from the center outwards.
Besides this, there also axonic connections between these structures, that are indeed shown in the illustrations, see e.g. above. These connections are probably feedback signals. So what one sees in the illustrations are the connections of the feedback, and not the forward ones, giving completely erroneous suggestions about the mutual relations between the structures.
Using this as an additional argument, we are going to include a structure in the basal ganglia, that usually isn't, see the illustration below (from the presentation here
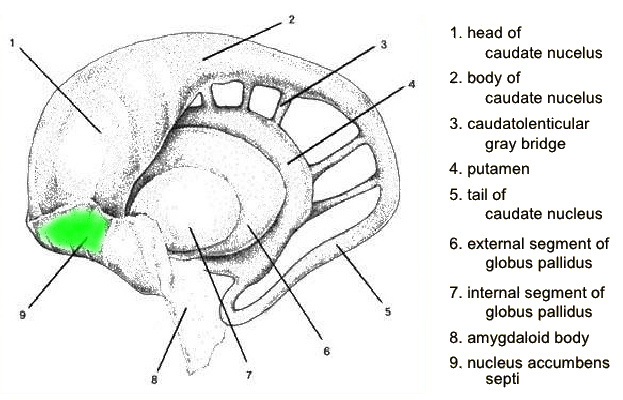 |
Highlighted in green and directly or neighbourly connected to the head of the caudate nucleus is the probably best known of the structures mentioned so far: the nucleus accumbens. Best known because it is known as the "pleasure center" and the one that regulates the release of dopamin, as we have seen among others the cause of addictive behaviour.
If one sees the basal ganglia as the next layer in decision-making, then the inclusion of the nucleus accumbens is quite logical.
This also corresponds to the alternative name given to the nucleus accumbens in specialized literature, as the ventral striatum (ventral = "on the side of the belly", in standing humans: towords the front), i.e. the more frontal part of the striatum where the dorsal striatum or putamen plus caudate nucleus is the backwards part. Denoting that putamen, caudate nucleus and nucleus accumbens have a common internal structure (the alternative one is the "pallidum" of globus pallidus and some other structures).
So including the accumbens into the basal ganglia is a logical step.
And the inclusion of the nucleus accumbens immediately (logically) calls for another one, i.e. its counterpart: the center of fear. This is almost as well-known: the amygdala. Now the amygdala is not neighbourly attached to the basal ganglia. But it is connected to them, though this connection is dropped in large number of illustrations. Here is one that doesn't, though it had be edited to show it clearly (adapted from here
 |
It shows that the tail of the caudate nucleus ends in the amygdala. This tale contains, of course, a bundle of axons.
And now we have all the ingredients of a substructure that can be described as: "a next layer in decision-making". Or: "a next layer in behaviour".
So what about the rest of the structures in the collection known as "emotion organs"?
Funnily, there is much more known about them than about the basal ganglia.
To understand why there indeed is yet another layer and why this is better understood than the basal ganglia, one further important point about the basal ganglia has to be mentioned: the fact that their internal connections, neuron to neuron, are almost exclusively of the blocking or inhibitory kind. Or in specialized terms: the neurons are almost exclusively of the "GABAergic" type.
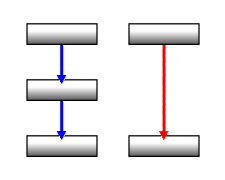 There they
may seems to be quite useless, but in fact this is not an uncommon situation
in logic: all logic operations can be constructed from two basic ones, but
both of them contain a negation, the logic equivalent of blocking - this is
because
There they
may seems to be quite useless, but in fact this is not an uncommon situation
in logic: all logic operations can be constructed from two basic ones, but
both of them contain a negation, the logic equivalent of blocking - this is
because -1 * -1 = +1 (on the right: blue arrows are inhibiting, red activating) - this doesn't work the other way around.
The passiveness of the basal ganglia may thereby denote they are a kind of filtering or memory system. It then just has to get its signals from elsewhere, a common technical situation.
So with this thought in mind, lets have a look at the other emotion organs, the ones about which much more is known.
Emotion organs, anatomy
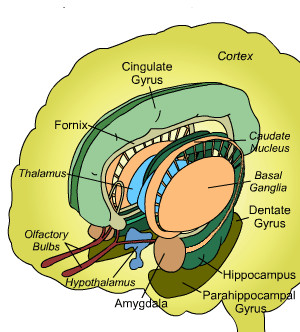
In the context of the basal ganglia the "emotions" are rarely discussed or even mentioned - in the literature one mainly talks about its role in movement. So what should be the real "emotion organs" must be located further on. Here is an overview of a number of structures beyond the basal ganglia:
 |
This is the same general area as that of the basal ganglia, and if depicted they would lie in the middle, but have been omitted for the sake of clarity. What you see in the drawing are structures surrounding them.
The central elements in this drawing are the amygdala and hippocampal formation. In our view of the organization the amygdala belongs to the basal ganglia, but this doesn't prohibit its appearance here: it may have taken on additional roles.
The nucleus accumbens is missing in the above and most other similar overviews, which corresponds to the choices made in this article.
The hippocampus is getting as well-known outside neurology as amygdala and accumbens, as a central element in the context of memory. More on that later.
Another important anatomical division between these areas is the anterior commissure and the lamina terminalis that is attached to it. The anterior commissure is a bundle of fibres going from left to right hemisphere of the cortex, perpendicular to the plane of the drawing. The lamina terminalis, which means "endpoint layer" is a thin layer, so it hardly shows up in cross sections - it does in the more detailed following one. In the present context the term "anterior" means "in front of this layer" (for additional similar illustrations, see here
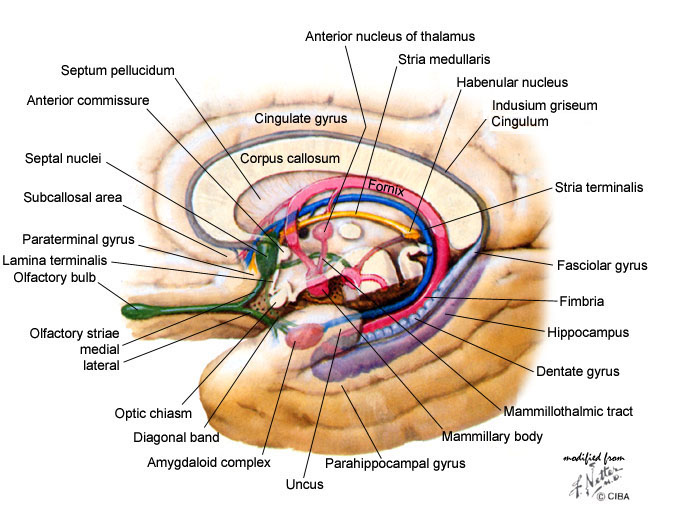 |
Also visible in this drawing are more nuclei and more connecting bundles. Their precise function, concluding from the several differing versions in the literature, is effectively unknown, with a few exceptions that be will seen later.
Note: in this second illustration is seen something that is usually omitted: the fornix has a branch in front of the anterior commissure, towards the (not shown) nucleus accumbens. And this also applies to the stria terminalis and the medullary stria, connections to or from other centers, which is also usually omitted.
The upper boundary of this all is formed by the fornix, which carries bundles of fibres from (and to) the hippocampal complex.
The second overview also tries to show another thing that is usually missed: the hippocampus is the boundary, going outwards sideways, between the area of the emotion organs and the cortex. More specifically: the sideways part of the cortex, at the front called entorhinal cortex ("inner side near the nose") and more towards the middle and backwards the temperal cortex ("near the temple") - see also the following vertical (coronal) cross-section through the middle of the thalamus (717 Gray - when comparing with Gray 742, please note that these kinds of cross sections can differ widely depending on the location at which you take them):
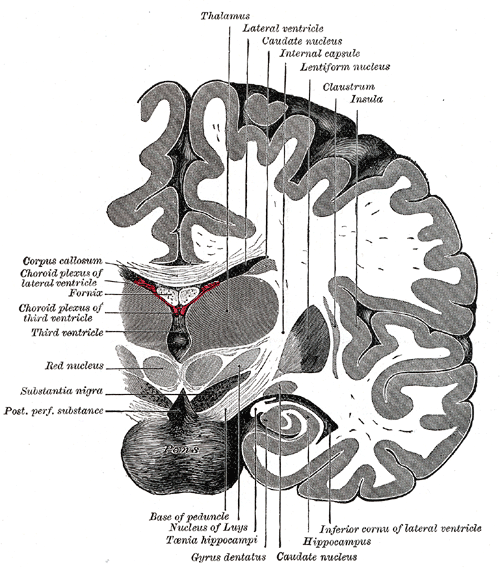 |
The curl denoted as "hippocampus" is in fact the parahippocampal gyrus (the first bend after the hippocampus proper) - the entorhinal cortex is below these two.
The windings on the right of the cortex proper, is the optical cortex - lightly indicated, by means of dashed lines, is the optic radiation, bundles coming from the brainstem and thalamus.
Some translations: Nucleus of Luys is another name for the subthalamic nucleus, and Taenia hippocampi is what is nowadays called the fimbria. The ventricles are canals filled with brain fluid.
This was the global overview of the structures in this area. To understand their more specific functions, it is of course first necessary to understand the function of the basic elements. This is: the role of these basic elements in that of the brain as a whole.
Emotion organs, function
As already seen, each new layer of functionality is there because it enables better decision making. While still retaining the previous layers, for application in situations that demand them - see this in action in this video
, where the old decision method (reflex: "flee for the lion ") fights for power
with the new (emotion: "protect the young"). One of the crucial methods for making better decisions is "learning". The list of components necessary for "learning" consists of: a memory, a recognizer, and an evaluator. Where the crux lays in the word "recognize". Which is a combination of "cognize" and "re-".
The "cognize" part means things like: "making sense of" and similar synonyms, which all boils down to: "You see something new that you associate with things you have seen before".
Or: learning is all about memory.
So where are these things to be found? Or probably: which of the already introduced structures is doing what?
The first step is standard knowledge, but applied in a novel way. That knowledge comes from a patient, known as "patient HM", by whom were removed both the hippocampi, in an attempt to cure him of severe epileptic seizures
Conclusion number one: the hippocampus is involved in making memories, but is not the (physiological) memory itself.
Conclusion number two: there are at least two types of memory.
The latter was already known
So this leads to conclusion number three: there are separate structures for declarative memory and non-declarative memory.
The last conclusion is supported by more evidence. From pathology there is the phenomenon of the Korsakov's syndrome
A second source of information also lies pathology: the cases of the damage in these areas as a consequences of oxygen deficiency, with as a frequent cause damage to the anterior communicating artery (ACoAr)
So the first question is: which parts are for which kind of memory?
And so the storyline returns to the jungle of physiology and anatomy.
One would expect that two systems of memory and associated systems would be clearly visible. Just as in the brainstem one would expect a step-by-step approach, with a sequence of structured performing each of the separate steps. And visible connections between them.
Now there is a series of connections has traditionally been noticed, after its discoverer called Papez-circuit
 |
 |
The "limbic system" is in the standard literature the equivalent of what is here called "emotion organs", with some differences, of which the major one is that the cingulate cortex, the first layer of the cortex, is included in the limbic system - as is done in both illustrations.
If one chooses its designation as Papez circuit, one gets the following sequence of elements
| hippocampal formation (subiculum) fornix → mammillary bodies → mammillothalamic tract → anterior thalamic nucleus → cingulum → entorhinal cortex, hippocampal formation. |
For the location of most of these components, see the above. Normally not shown or mentioned in this context is the cingulum - this a collection of structures in the area lying between cingulate cortex and corpus callosum, also referred to, also referred to as the indusium griseum, denoting it contains neuron bodies (Gray 751):
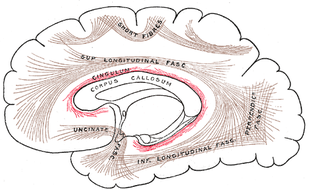 |
But the cingulum also contains axonic bundles that connect several structures, among which, as shown, is also the hippocampal complex (at the lower end; not named). The connection from thalamus to cingulum is not shown, but this is part of the much larger bundle of connections between the brain stem and thalamus and cortex seen earlier: the internal capsule.
Abbreviated the circuit now becomes:
| hippocampus → fornix → mammillary bodies → mammillothalamic tract → thalamus → cingulum (entorhinal cortex →) hippocampus. |
Herein, the thalamus, the structure is the closest to the brainstem, and those in which presumably the virtual reality of the brain is first constructed. So this is the logical starting point for the process of the formation of memory - and because the entire track is circular, you can simply shift the starting point, giving ...:
| thalamus → cingulum (entorhinal cortex →) hippocampus, fornix → mammillary bodies mammillothalamic contract → thalamus |
... having the format: structure 1 - connection 1 - structure 2 - connection 2 - structure 3 - connection 3 - structure 1.
Of the second structure in this loop, the hippocampus, we already know the role as an essential element in the formation of declarative memory, and the effect of damage to it results in anterograde amnesia.
But anterograde amnesia also occurs when suffering from the syndrome of Korsakoff's
In other words: the Papez circuit is that of formation of new declarative memory.
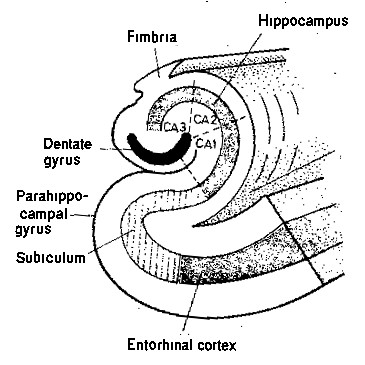
But what is the specific role of the hippocampus in the formation of memory? A first indication can be found in its overall shape, visible in schematic illustrations such as these (from here
 |
This tapered shape makes a technician think of a sort of filter function, with the final product discharged from the tail (the fornix) to other nuclei (the small globules at its end are the mammilary bodies).
At this point pathology again comes to the rescue. During operations on another patient suffering from severe epilepsy, one could perform experiments with instruments so fine that one could follow the activity of individual neurons. The first of these experiments became famous for the discovery of the "Jennifer Aniston"-neuron
Later experiments showed that in fact there were lots of other neurons that also fired when such a picture or similar ones were shown, and that in fact what fired was a sequence of neurons according to abstract concepts, like object - mover - human being - male / female - ... etcetera
And this obviously is a branching process: if the individual is "male", there follows a different sequence then if the identification is "female". If the person is a male, the female alternative is something like this: ... young - blond - breasts - ... and so on.
Very "sexist", all this. But the alternative with the roles reversed is equally so: long - heavy - athletic - ... and so on. Also very sexist, but from the viewpoint of "security" and "social position" instead of "beauty" which in fact is a measure for "getting healthy offspring".
Back to the neurology: the tapered shape of the hippocampus corresponds to the increasingly more abstract concepts, of which there are less thus there is less to filter in the next stage so thois can be smaller. With the highest abstractions to be found in the tip of the tail.
Incidentally, the survival or evolutionary value of developing such a system of abstractions lies in the advantage that a fish doesn't need every other fish it encounters to give a separate review, but that it can be classified using specific general characteristics, thereby coming quicker to the desired result: "Predator or not?".
So now we know about the overall circuit of declarative memory, and about the analyzing or filtering function, but not yet about the memory itself. Further information on this can be gained from the more detailed structure of the hippocampus and surroundings. A first glance was visible in the cross section of Gray 717 above and in more detail here, which is a cross-section perpendicular to its length as shown in the previous illustration (from here
 |
This clearly is a "from inside to outside" structure (also immediately clear is why the hippocampus has its name: it has the outward shape of a sea horse or hippocampus).
Now couple this with experience gained from everyday life. When someone gets involved in a sudden traffic accident, it is a not uncommon phenomenon that he or she cannot remember anything of what went on directly before. In fact, this period of memory-loss can be specified quite precisely: it is about twenty minutes.
Add to this: in the context of education, research has been done on the attention span of children at secondary level: its maximum is about twenty minutes (any able teacher can confirm this number).
So these two findings can be combined into a picture: the pipeline of direct sensory information, through primal filtering and interpretation, through the hippocampal analyzer, through the process of adding other information, takes all in all about twenty minutes. And only then the permanent memory has been reached.
The permanent memory for experiences that run through the hippocampal circuit is, as the picture shows, the neocortex.
This solves one half of the puzzle posed by that patient HM and similar cases: the loss of new declarative memory (by the way: he retains his old memories because he retains his cortex!).
Still remaining is the other half: why doesn't it effect the episodical or procedural part of life, the daily routines, and where does that fit into the neurological picture?
This is the next subject that starts with an inventory of what is already known of the various types of memory.
Memory
In standard neurological literature there are several types for memory, with the main division being that already introduced: that of declarative and non-declarative. However, its previous use here doesn't complete match the usual way this division is made, so here is an overview of the various known types of memory, where the definitions as given in Wikipedia are followed, because in most cases interested people will pass through there anyway:| Name | Subname | Content | ||||
| declarative, or explicitly |
episodically | observations belonging to firsthand experience or nearby - with a sense of time. Especially in combinations. | ||||
| semantic | knowledge - facts, ideas, opinions, concepts. | |||||
|
non-declarative, or implied, or procedural |
unconscious action, unconscious behaviour. Also in combinations. |
Read the more elaborate descriptions in the original sources, and get an idea of the confusion.
So here new divisions are introduced. With one question up front: new names or new definitions? The choice has gone to a mix, in order to get a result as close as possible to intuitive notions.
One thing is clear from the descriptions given so far: "procedural" and "episodic" belong together. Similarly: the semantic memory is the one involved in the hippocampus.
Before knowing the precise function of the hippocampus, "declarative" seemed a suitable name for its output, but afterwards "conceptual memory" or "abstract memory" seem better choices - "declarative" is retained as a synonym.
For the form that runs the daily affairs, the choice has gone to "episodic memory" because it is more specific than "procedural".
Together this largely automatically coincides with the intuitive distinction between conscious and unconscious: patient HM lost the capacity for storage of conceptual memory and conscious memories, but could still learn episodic or procedural or unconscious tasks.
But now we are still left with the problem: where does the memory for the episodic part of life reside? So back to its physiology: the basal ganglia.
Basal ganglia, part two
The declarative or conceptual memory was treated before the non-declarative or episodic not because it is more important or more primitive, but because it is "pronounceable", i.e. accompanied by verbal expressions, i.e. much easier to follow when something goes wrong due to physiological damage. It is the other way around: damage to the episodical system has much more severe consequences and quickly leads to death, from which little can be learned.But with the acquired knowledge about the declarative system and the suggestion that there is probably still some similarity, it is now possible to trace the episodic system much more easier.
First: assume it is also a circuit with several components. Secondly, assume there is also a kind of filter to separate the important from it with lesser. Thirdly, from the fact that patient HM could still lead a fairly normal life, conclude that the episodic memory is more primitive and more essential, so that its structures are probably located closer to the brainstem. Together this shows that the answer has already been given: the system of the episodic memory is that of the basal ganglia.
So we again have a walk through them.
The logical starting point is the thalamus, the relay center between brainstem to the rest of the brain. Already mentioned is that it has a connection to the basal ganglia coming from its centromedian nucleus in its core:
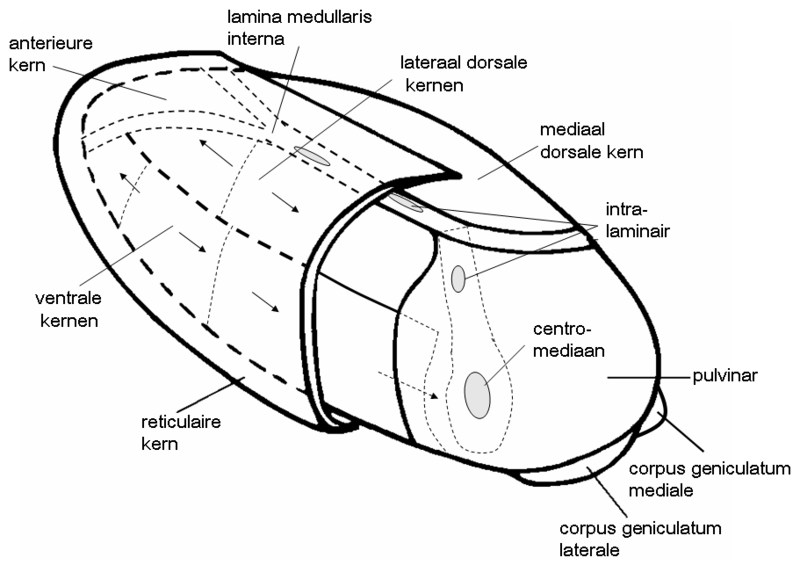 |
So probably it relays all the sensory and related information gathered in the thalamus to the basal ganglia, more specifically: the globus pallidus interna, the innermost of the series of the basal ganglia.
Remarkably, on what goes on next, information can be found in standard literature, in a different and more isolated context. Here is an illustration representative of this story - it is a horizontal cross-section at the level of the thalamus (from here
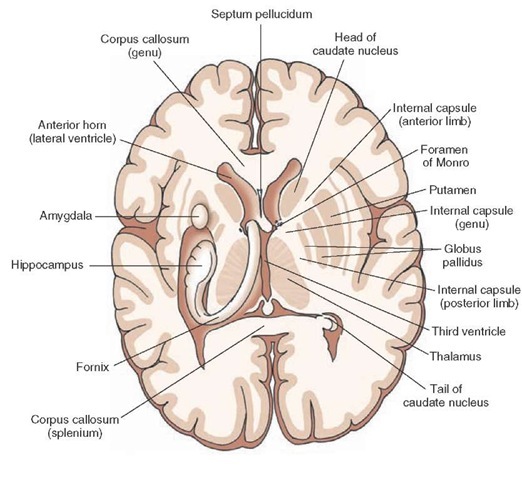 |
This is about the right hand half of the picture, where only one interpretation makes sense: the information or control flows from inside to outside from the thalamus to internal globus pallidus to external globus pallidus to putamen. And in some of these stories one also goes beyond the realm of the emotion organs: to the claustrum (the next very thin unnamed layer) to the insula (the unnamed C-like protruding portion of the cortex).
And in this way it is often presented at academic introductions in neurology for students.
So this is the general flow, corresponding to the sequence of structures in the hippocampal complex.
The next step is to find the part that does the filtering. Remembering one argument that was used to identify the hippocampus: its tapered form, there immediately rises one candidate: the caudate nucleus.
So lets repeat our earlier illustration, that was chosen because it shows, besides the connection to the amygdala, one other thing that is frequently omitted: the series of bridges between the tail of the caudate and the putamen:
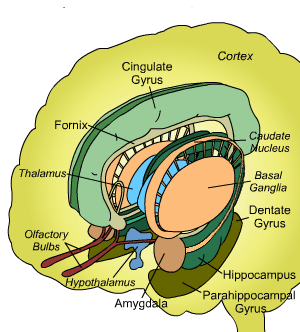 |
Now when thinking like a technician, you immediately associate this kind of structure with a filter that has tapping points at regular intervals.
With an added property when you realize that the caudate is part of an information flow from inside to outside: the bridges are points where information is fed back - where output is fed back to input. (By the way: it might also be a forward feed, but later more arguments for the former will follow)
So clearly the caudate nucleus has a filtering and regulating function.
With its final output going to where things go that matter the most: the general warning and alarm system of the brain: the amygdala.
Explaining the already mentioned fact that the caudate nucleus and the striatum in general almost exclusively exists of inhibitory, blocking, non-pro-active neurons.
But if the striatum is, like the hippocampal circuit, a recognition and assessment center, there remains the question: where is the memory itself?
But again we can use the analogy: is there something that looks like what follows the hippocampus as its memory system which in that case is the neocortex?
This again is a case of "posing the right question", because the answer follows immediately (if one has previous neurological knowledge): this is the cingulate cortex, see the illustration below where the cingulate cortex is yellow and the neocortex is green:
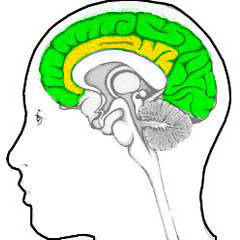 |
The big thing (literally and figuratively) that distinguishes humans from apes and monkeys and has grwon so much, is not the cingulate cortex, but the neocortex. So when one uses the term "cortex", what one usually means is "neocortex" (see e.g. the illustration before the last).
This difference is also reflected in the internal structures: the cingulate cortex has five layers of neurons, and the neocortex has six. That is: the neocortex has an extra layer for extra functionality.
Now combine this with experiences from everyday life where people appear to perform whole series of actions "on autopilot". You start an activity, and doesn't notice any further steps, until you get notified that it is finished. Which is quite handy, because you can turn your attention to other activities, like thinking about what you have to next when it has finished. And think about what happens to for example a piano player when he starts think about how to move his middle finger, that is, turning on the hippocampal circuit: he gets hopelessly confused with his routines that constitute of fixed combinations of fingermovements.
So this seams to be the major difference between the episodical circuit in the brain and the hippocampal: the former starts and then winds itself down, the latter can be interrupted at will. With corresponding experiences from the memory part of things: it is very difficult to nearly impossible to remember the intermediate steps of an episodic activity, and it is relatively easy to do this for hippocampal, declarative, memory (or the other way around: that is why it is declarative!).
And therefore the extra layer in the neocortex!
Now one more riddle in the puzzle remains: how can this have been missed?
It wasn't, really. It just wasn't noticed. Here is the limbic system again:
|
|
|
Including the cingulate cortex. And here the associated pathway:
| thalamus → cingulum (entorhinal cortex →) hippocampus, fornix → mammillary bodies → mammillothalamic tract → thalamus |
With the cingulum. And here is the cingulum:
|
|
So there is a direct path from thalamus to cingulate cortex, that is: it is more direct than to the hippocampus.
This doesn't preclude that there is also a path to the hippocampus, but it might not be the primary one.
So this is the result so far: there are two ways to deal with reality: a primary one that does it by way of fixed scenario's, and one that "analyzes" reality in groupings of abstract concepts. Or in everyday language: the way of "acting" and the way of "thinking". Old knowledge, of course, but now translated into neurological structures.
Whereby the "acting" part of things can be readily imitated, as already found out long before the advent of the computer through the "automaton"
 |
 |
Where even the mechanical "automaton" on the left can behave eerily human looking (when suitably dressed), while it just follows a fixed list of instructions coded in paper, stored in a way like in the illustration on the right.
Now the big difference between the automaton and human behaviour is that the latter has to be learned. Which goes both for the acting and thinking part of it. But before going over to that, there is one more aspect of the acting things of the brain that has to be discussed: its role in dealing with emergencies.
As things stand now, this is still dealt with by the brainstem, where the neurotransmitters to put the system in a state of alarm and back again, with a controlling system that does it in a rather abrupt, "on" and "off" way.
The basal ganglia also have improve on that.
Already mentioned and given some importance are the bridges between the tail of the caudate nucleus and the putamen. These are probably a form of feedback but could also be forward coupling. But in both cases it could also be used to speed up things: by shortening the response time of the system if you take that the signal running through the entire tail of the caudate is the standard.
So the caudate facilitates a more gradual response to situations in reality that tend to being an emergency, saving energy and resources.
For this to make sense, it has to correspond to the experiences of everyday life.
That it does. Look at your own life.
Besides that, everyday life adds another experience that until now didn't have a place: during an emergency the sense of time changes. And rather drastically at that. From the time that instrumentation was not as developed as it is now comes a story about the experiences of test pilots. Of course the engineers wanted to know "How long did the emergency last?", and test pilots with all their flying experience and being the most cold-blooded ones around, gave answers as well as they could. As soon as one was able to record the entire flight, it turned out that the test pilots were off by a factor of three or more.
This can be build into the model, introducing what is seemingly the first entirely unsubstantiated assumption into the model: take that the processing of observations of reality is not a continuous process, but comes in a sequence of distinct frames, just like a movie (18 frames per second) or a television (25 or 30 frames per second):
 |
Everyone sees this as a continuous flow, so if in the brain something similar happens, one gets the same continuous impression.
And once this idea is posted, there are several observations that seem to point in this direction.
- There is the technical argument: a continuous series of images, or any other continuous (analog) information, it is difficult to analyze and keep in control. This is much easier to do on a separate frame, that remains the same for the time of the frame rate. For example: the nuclai in the brainstem must be activated by a sigal in the emotion organs, and signals take time. And production has to start. Etcetera. The modulating signals are by definition somewhat later than the observational data, and would run out-of-sync when the signal was continuous.
- There is the pathological argument: the most common neurological disease is epilepsy, in its various forms. Epilepsy can be induced in a certain group of otherwise normal people by rapidly successive flashes of light - British television warns viewers for "flash photography". Physicists know this phenomenon as "resonance", if there is a process in the brain that also works in series.
And the frequencies at with induced epilepsy phenomenon occurs, lay in the range of 10 to 20 and 30 per second, the right range.
- There is the phenomenon of the "dead time zone": when things happen or change within a certain time frame, they are unnoticed, when the time frame is short enough. With many familiar examples. Card magicians and the like use this by "being quicker than the human eye can follow".
Another example: when designing the human interface for a computer program, the changes must take longer than a certain time frame, or else they get initially unnoticed especially when the changes are small - and to get the impression of a continuous transition, you must use a minimum time frame for this transition. Which designers know very well: it is about 0.12 seconds. And since there must be at least three picture frames in this period, one can estimate the associated frame rate: around 25 pictures per second. Right in the middle of the earlier given range.
- And already noted is the time-stretching phenomenon: take that the caudate nucleus is the one that determines the frame rate, where the standard rate corresponds to the signal running though the entire tail. Then a shortcut though one of the bridges raises the frame rate. And the frame rate determines the experience of time, since there is no objective of measure in time to be found anywhere in the universe.
So the entire process at this point is: the caudate nucleus filters the observational data and compares this to known patterns. When a hit occurs against a type that has to be avoided, it short circuits the regular loop increasing the rate of observation and the rate of action, and sends an alarm towards the amygdala. Etcetera.
Everything one expects of a mechanism dedicated to survival.
At the location where one expects it: as soon after the brainstem as is possible. And almost continuously adjustable, in sted of just "on" or "off".
The hippocampal circuit is probably directly coupled to this frame rate, but, as again plenty of everyday experience proofs, feels and acts like an innocent bystander.
At which point it is time to notice that in an area that is denoted as that of the "emotion organs", little has passed by way of "emotion" in the usual everyday life meaning of the word. So for the moment we pass on another "mechanical" thing that has been mentioned but not addressed: learning. And go over to some real emotions.
Higher emotions
So far the emotion organs have shown little of dealing with emotions, if one takes out the more reflex-like responses of fear and a kind of stimulus that invites to addiction. And please note: much of the previous structures and processes can also be found in reptiles.But if the extremes of these urges aren't touched, the much more finely adjusted control of behaviour and thereby observation of the surrounding world gives room for more subtle things.
The likely next step would to take more care of the offspring. Dropping eggs in a hollow and then pouring sperm over it, as fish do, might work under water, but in the ever changing environments abvove it, more care of the offpsoirn is highly advantageous: the new generations don't have to learn what is effective behaviour all over again, but learn it from the preious ones. In humans this is called "culture", a term that used to reserved for humans but of course applies to all mammals.
When expanded to wider circles, this leads to more evolved forms of group behaviour. Living in groups also has great advantages: there are more eyes so earlier warnings of danger, more noses, so rather the smell of food. Individual weaknesses and strengths can compensate each other: the strong male wildebeest can defend the young helpless wildebeest against the lion. In a further phase, the sharing of food: the individual keeps foods available not just for themselves, but partly leave this to a group member who is more served with it. So it survives and later can help you, so that the net sum for both is positive.
But these are all things that have to be build upon the already working things, so for example the mechanism that controls care of offspring and group behaviour still use the same tools as the brainstem and the basal ganglia: neurotransmitters.
In the area here denoted as that of the "emotion organs", lie the structures that make these new neurotransmitters and regulate them.
These neurotransmitters are nowadays, writing 2017, well-known, but some twenty years ago this wasn't the case. The best known is oxytocin, first given the popular name of "love hormone" because it stimulates motherly care. And later on it was joined by vasopressine, the neurotransmitter that was proven to be involved in male-female bonding. With other ones they can be called "tertiary neurotransmitters", with glutamate and GABA being the first layer, and norepinephrine etc. being the second. These tertiary neurotransmitters are then the agents of the "real" i.e. commonly denoted as such "emotions", and their associated structures the "real" emotion organs. Which shall not be dealt with in detail, because even the neurotransmitters themselves have given rise to controversy.
First of all there is a the great unease among large groups to describe anything human through rational mechanisms. Which gets successively worse with every step that comes closer to human behaviour, and even more so for human thinking.
And oxytocin soon gave rise to qualms among the ranks of people who are not infected with these social viruses, when it was discovered that oxytocin works on all kinds of group bindings, and not only in the positive sense, i.e. being active by binding a certain group of individuals together, but also in the negative way, by repulsing individuals that were not part of this group.
Now this is as natural as it can be, because the concept of "group" logically includes the concept of members, and by the same token the concept of non-members, and when expressed in active terms one then gets the same pair, but then as "attraction" and "repulsion".
Nevertheless this gave problem is some quarters, besides the expected ones of Christians who believe in the non-existence of enemies and "turning the other cheek" etcetera. But also among non-Christians there are large groups that believe that humankind is destined to be one "happy family" under the uniting rainbow, at which end we will find the pot of gold once this noble goal has been achieved.
And the existence of a neurotransmitter that besides doing the bonding also does some rejecting or repulsing, is a thought quite repulsive to these people.
And with vasopresssine
Now this comes already pretty close to all kinds of theories pejoratively denoted by "isms" that are absolutely forbidden because of previous misuse. So we stop the description of the emotions here besides some further remarks made in the section Neurology → sociology.
Learning
So finally we have arrived at the point where, again according to large groups of people, the boundary between animals and mankind lies: the faculty of learning. Of course, there again one is wrong, as with almost every other statement of this kind, because all living creatures have to deal with changing environments, and those that don't learn in some kind or fashion, simply don't exist anymore. In America, one has started shooting bears again, in order to unlearn them the recently novel experience that they weren't shot by humans, and that the human environment had richer pickings for them than the outbush.This learning from the environment is done by the same structure that do the filtering and comparing, and as far as the analytic part of the emotion organs is concerned this is the hippocampus - and large swathes of living creatures have a hippocampus (two, to be precise). But how is this done?
Now both in nature and in human technological endeavour one has discovered a kind of structure that is able to do learning: the neural network. A first application of such a technical network was the recognition of handwriting on bank statements. Such a network is trained by giving them lots of samples of human handwriting, with the corresponding correct answers. The network learns itself what to look for.
All conglomerates of neurons could be called "a neural network", but what one specifically means is "a layered neural network". This is: a network that consists of layers of neurons with layers of connections in between.
Which immediately provides the definition of the simplest neural network, but in order to stay with the familiar, one step further is taken and gone to the next level, since the neurological part of the eye, the retina, is a three-layered neural network. Here is a real-life and a schematic representation of it:
 |
 |
The neural network of the eye does relatively simple things like translating the picture of the outside world into contours, lines, and fields, area's, and send these concepts to the brainstem. Instead of the complete continuous and "analog" picture.
This is thereby a simple form of what the hippocampus does: extracting distinct abstract concepts from a continuous reality. Here is a closer look at the hippocampus, more precisely: the head of it:
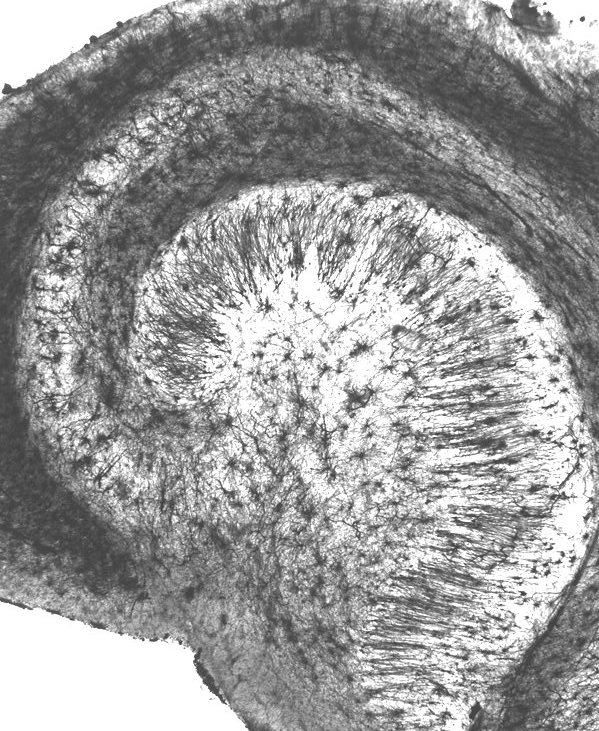 |
This clearly also is a layered neural network, though because of its length rolled up in an compact curled form, and having more layers.
Now the whole point of such a network is that it can be trained, as is known from the technological version. It does this by strengthening (activating) certain internal connection, and weakening (inhibiting) others. A procedure that is too inolved to treat here.
However, what is known and must be discussed is that the training of such a network is in fact done not in one, but in two stages (note this recurring theme): training on "false negatives" and training on "false positives", or "Finding out what does apply," and "Finding out what does not apply."
Now "training on false negatives" is similar to recognizing things, so may at least partly be done concurrently with the normal everyday life activities. Which cannot be disrupted on too long a basis, so learning during the day remains limited.
For the other phase, training on false positives, this is not an option. To train on false positives, one needs input that does not correspond to reality. And it would be very disturbing to the individual if he would during everyday life and on a regular basis would be confronted with imagery that is false and distorted.
At which point one can readily guess when this can be done: during sleep. Here is an example of the brain activity measured (by an EEG or electro-encephalogram
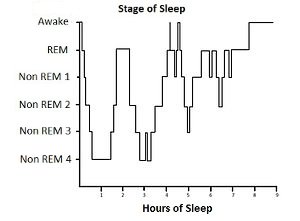 |
Showing two main types of sleep, called "deep sleep" and "REM sleep", and the latter because of the rapid movements of the closed eye one can observe. It is also the time when people have dreams, proven by wakening them.
Of dreams it is overly familiar what their main characteristic is: they show unrealistic to absurd distortions of reality. Things that do not have happened. Precisely the things that are needed for the training through false positives.
So the block-like shape of the sleep-cycle has now an easy explanation: they are successive cycles of "false positive"- and "false negative"-training. Just what would expect from an efficient cycle: first gather a lot of positive identifications, then weed out the optimistic ones, and with that experience gained, again take a batch of positive identifications, and weed this one out. Etcetera.
But non-existing images are what they say: they are non-existent. So they have to be made. By the brain through a neurological structure. So the question is: is there a suitable candidate for this task? So lets have another look at the area of the emotion organs:
|
|
On the bottom of this inner area there is a part colored in dark-brown, a very unevenly structured field with a function so obscure that the area wasn't given a name for a considerable time during the first wave of anatomy on the brain, after which it was given the name of substantia innominata which is Latin for "substance without a name". Very appropriately, it also is unnamed in the illustration above.
The substantia innominata fulfils both requirements for our structure that makes unreal imagery: it has no known function, and it has a very irregular internal structure.
Here is an illustration in which the substantia innominata indeed was given its name (from here
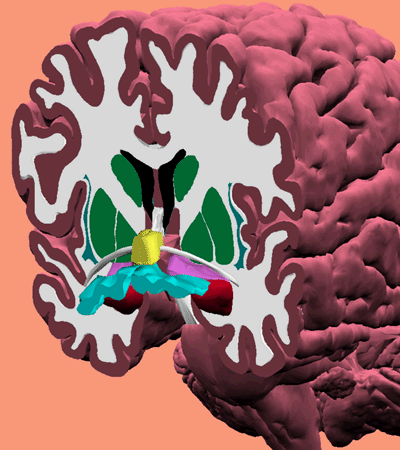 |
This is a (rare) frontal cross-section, at the location of the basal forebrain - the top green area is a cross-section of the familiar caudate nucleus (here right and left version), the green below it is the combination of putamen and globus pallidus. New things are the yellow ones (the septal nuclei, part of the reward system), the white beam passing by them is the anterior commissure (the frontal connection between the left and right hemispheres of the cortex), purple is called the basal nucleus region, red the familiar amygdala, and blue (at the right side partially cut away): the substantia innominata.
In addition to these anatomical clues, there are also functional ones, as before from the field of neuropathology. A common source of brain disorders are strokes, and specifically for this functionality those to a local artery: anterior communicating artery (ACoA). One has recorded two main effects of these strokes, as described here (from: memorylossonline.com
| The patients that DeLuca works with confabulate because of a rupture in a tiny blood vessel in the brain called the anterior communicating artery (ACoA). The rupture of this tiny artery temporarily cuts off the normal flow of oxygenated blood to areas of the brain that are essential to the proper recall of memories. The damage caused by an ACoA rupture can vary from one person to another, both in the location and the degree of damage. And the symptoms are also diverse: the person can suffer memory impairment alone, or memory impairment accompanied by confabulation. |
The loss of memory shows that the affected areas are part of the stream of information of the conceptual or declarative memory:
| For reasons not entirely clear, damage to the basal forebrain can impair the ability to form lasting memories from recent experiences ... |
The basal forebrain is the area we're talking about here.
So this study reports two phenomena: anterograde amnesia and confabulation. The anterograde amnesia is the one been mentioned before as an indication that it belongs to the circuit of the declarative or conceptual memory.
Confabulating
Add this to it: the phenomenon of confabulation doesn't just occur after ACoA-infarct, but also in combination with Korsakov-related damage.
All taken together this suggests that there is a structure within the area of the emotion organs whose task it is to chop and distort experiences in all kinds of ways. A structure that is intended for use during sleep, but by various forms of damage to it or its surroundings, can also become active during the waking state of the mind. At which point the regular route of evaluation via the hippocampus is then (partly) turned off.
This kind of disruption of normal operations is familiar from a wide range of neuropathological phenomena. The people suffering from them all report particularly peculiar sensations. Which for them are as real at that moment as are actual sensations, because the actual sensations also are nothing more than an information flow in the brain. The best known of these phenomena are also the severest: psychosis and schizophrenia.
Concluding this section: the hippocampus seems to have two major functions: filtering and analyzing during the time of wake, and getting trained during the time of sleep. This involves different flows of information, e.g. to and from the memory. At least a part of the structures in the area of the emotion organs will devoted to this task. Which has little to do with "emotions" in the everyday-life sense of the word.
Overall structure
So the emotion organs in fact comprise of (at least) two main sections: the basal ganglia doing the everyday routines, and the hippocampal circuit, doing the analyzing etcetera. This comes above the structures of the brainstem and spinal cord, that do the basic tasks of observation, motion, and housekeeping.These things all have to work smoothly together, of course, for them to maintain their evolutionary advantage. As the better human engineers do, nature does this not by a straightforward one to two to three model, but through a number of feedback loops. System 2 one sends signals to signal 1, and system 1 reports back on what it has done, and system 2 adjusts its further actions. Etcetera. Normally the emotion organs tell the brainstem what to do. If there is an emergency, the brainstem takes over and shuts out the emotion organs. Etcetera.
The following illustration belongs to an explantion of what goes wrong when someone is suffering from Parkinson's disease (original from here
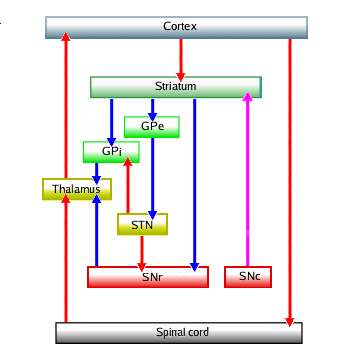 |
The circuit shows a number of the loops that control movement, starting with the intention to in the cortex. Red arrows are the activating connections, blue the inhibiting. They are modulated by dopamin coming from the SNc: the substantia nigra in the brainstem. Not shown in this original circuit is the counteracting one, acetylcholin, is not shown.
Everyone knows the major symptom of Parkinson: a tremor of e.g. the hands. And every engineer knows what causes oscillations, or tremors, in a system: something going wrong in a equilibrium system controlled by a feedback loop.
What does go wrong in the case of Parkinson's disease is a loss of the ability to produce (a sufficient amount of) dopamin, causing under-stimulation of one part of the loops and thereby over-stimulation of another - causing the shown loops to oscillate. Here is the associated illustration of the circuits in a more life-like fashion (from here
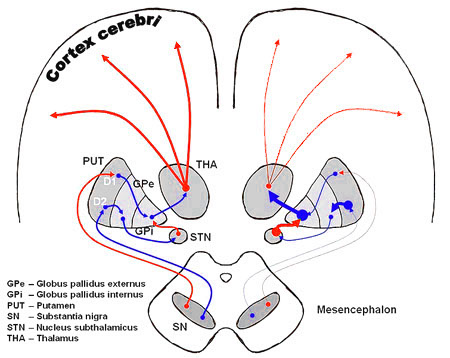 |
Visible are both hemispheres, with on the left hand side the normal situation of a healthy person where everything is balanced. The right-hand side corresponds to a deficiency in dopamin, and leads to the tremors: vibrations in movement due to disturbance of the feedback loops.
There usually is one thing missing in these kind of diagrams, and that is the distinction between the main flow and the control, see the archetypal example of a steam engine (from here
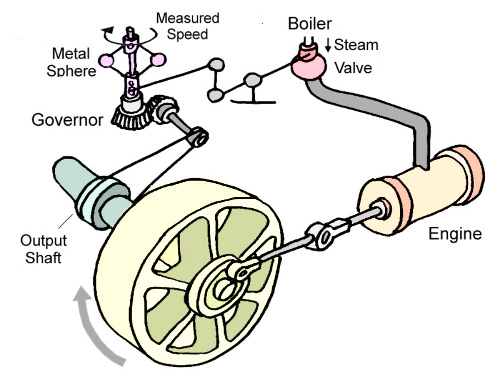 |
The main power source is the energy stored in the steam coming from the boiler, in the cylinder (the "engine") passing over to piston (not shown), thick connecting rod, to the flywheel - then the control circuit starts with a thin string from the output shaft to the "governor" that determines the speed and adjusts through some thin levers connected to the steam valve the amount of steam going to the engine.
All these or equivalent elements must be part of each feedback system, so also those of the neurological versions, and without having them all in the picture, the task of understanding the whole is nearly impossible.
It is highly probable that most of the yet unmentioned structures falling under the collective name of "emotion organs" are of the physically smaller type of "controlling function".
The Parkinson scheme lends itself for some steps in this direction, since it involves mainly or exclusively the episodic system. Here is such an extended scheme:
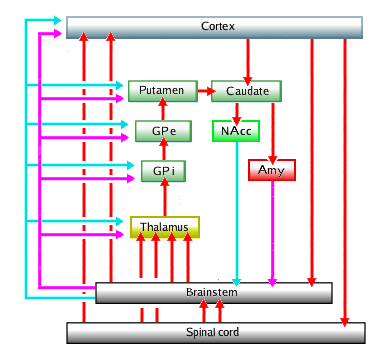 |
Immediately clear is the value of including the nucleus accumbens and amygdala into the basal ganglia: one gets completed feedback loops, with the main, "engine", parts denoted by the red arrows standing for the main information stream (through glutamate and GABA neurons), and the control or modulating circuit denoted by the magenta and cyan arrows standing for the dopaminic and acetylcholinic neurons (in the brainstem) and their neurotransmitters - for clarity the activating neuron lines from accumbens and amygdala towards brainstem haven been given their control-function color.
The intimate way in which the three main structures of the brain work together can also be seen in this very much simplified global connection scheme (Gray 764)
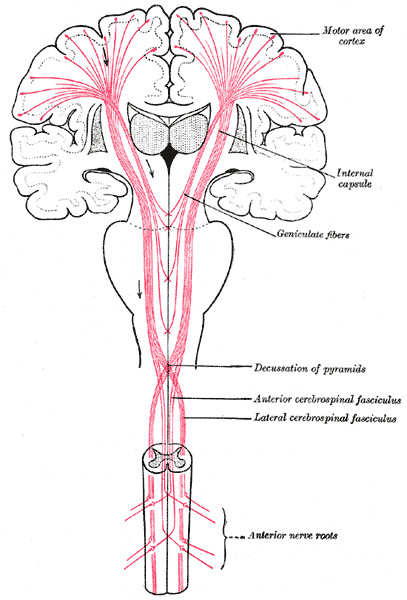 |
The gray globes near the top are the thalamus's and the gray triangles sideways of them are parts of the putamen and environment. The heart-shaped thing in the middle is the cerebellum that facilitates fine control. Both are also connected to this scheme, which is not shown (for clarity, probably). Drawn in the spinal cords are some nerves going to muscles etcetera.
And one more system of connections deserves some attention, because it appears with some regularity in standard treatments, as a continuation of the relations among the earlier mentioned axis: thalamus, globul pallidus and putamen, here in a repeat of the previously used image Gray 717:
|
|
The next element on this axis is the claustrum - literally: "cloister". Though very thin relative to the others, one supposes important roles or it, because it seems to connect the basal ganglia to the strongly protruding part of the neocortex, the insula - literally: "island". Comments are made that this conglomerate plays an important part in consciousness. But these comments still the have status of speculation. The functionalities added here to the basal ganglia, gives a bit more weight to them.
Rounding off
Here an attempt has been made to bring some order in the body of descriptions in neurological science that in fairly large extent still is somewhat of a mess. The central idea was the evolutionary approachThe main lesson of evolution in this respect is that it usually builds on existing structures, because the existence of these structures, in the longer term, proves that they function. And for new functionalities nature builds on these old ones, thereby creating chains of functionalities, of which that of spinal chord → emotion organs → cortex is an example.
Note that this absolutely does not have to be the most optimal solution - by designing something from the beginning to the end, a better functioning whole may be possible - the wheel is by far the most efficient method of locomotion, but nature could not develop it, because at some state you have to drop the use of legs, temporarily. And at that point, the living creature is deadly vulnerable.
Of course, there are explanations other than the model outlined in this article. However, the requirement to these other explanations is that they must lead to at least as much consistency between the different observed phenomena as the model outlined here.
There still follow some sections on the relation with other fields of knowledge.
Neurology → language
Of the related fields that are influenced by neurology one has already been frequently mentioned, though implicitly: that of linguistics. The very name of "declarative memory" says something out "speaking out", i.e. language. Which is closely related to thinking, a young child once heard remarking: "Thinking, that is talking in your head". Not exclusively, but true enough in many senses.Though the research that discovered the process of abstracting used images, there is every reason to believe that the recognizing of the meaning of language uses the same process. Not only the memory, but the whole process is "declarative". As demonstrated by the endless stream of words by people that boast some ability in the direction of "understanding" and "thinking".
The latter phenomenon, and especially the associated misuse, has not gone completely unnoticed, e.g. on the accompanying phenomenon of the use of words and language to manipulate other people. For example George Orwell used it in his novels, like 1984
The most consistent effort however has come from linguist Alfred Korzybski
 |
In which form it gained some notoriety, mainly in the field of creative writing.
Of course, what is depicted here is precisely what the neurologists involved in the Jennifer-Anniston-neuron research discovered.
From these discoveries on the working of abstractions in language, one could have worked the other way around, when realizing that much of ordinary behaviour has a basis in neurology. This is in fact is precisely what Korzybski attempted in his main work, Science and Sanity, but could make only little headway with, because of the lack of neurological knowledge at the time. This relation can now be described in much more detail which is done here
Neurology → Psychology
Of the main fields of psychology: pathology, public information, and scientific description, the isn't any that one could think of doing without an thorough introduction of neurology as for example given above.The only discussion one could have is about public information, but usually "the public" gets interested in these kind of things when they experience some "difficulty" of kind or another, and a very large proportion of these "difficulties" arise from the fact that there are indeed three main subsystems in the brain, and that at times they give off conflicting signals: a quite rare example where this has also been seriously researched is the behaviour of people that get involved in an airplane accident. A large group panics and endanger themselves unnecessarily, and a large group paralyzes and endanger themselves unnecessarily. How to prevent this?
The fact that these divisions exist has not gone completely unnoticed, of course. The difference between the smaller group of people that that to use or trust the rational i.e. declarative part of there mind more than the emotional one and the much larger group (20 to 80 or there about) for which it is the other way around is well-known. With similar divisions formulated as "analytic" or "rational" versus "holistic" or intuitive" etcetera. With both having their merits, otherwise nature had not developed this dual system which is in effect triple (together with the reflexes).
A surprisingly effective depiction of this state of affairs shows this illustration below (at several places on the internet, original source unknown)
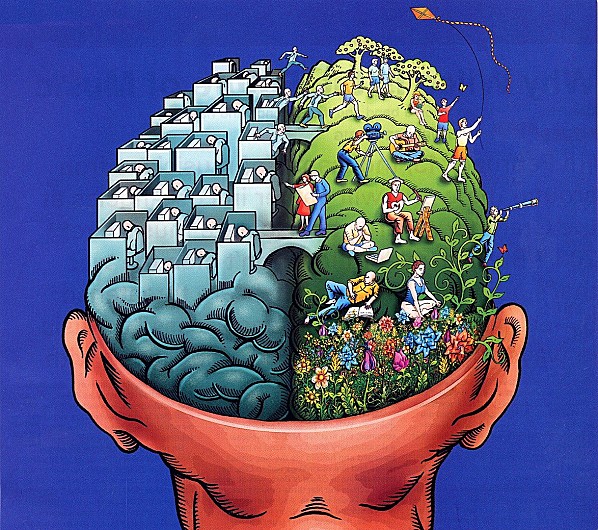 |
With the holistic or episodic approach executed by the basal ganglia in the right hand hemisphere, and the abstracting or analytic approach executed by the hippocampal system on the left hand side.
As is explained in more detail in the already mentioned article on the abstraction ladder and the hippocampus, this version looks like the brain of an infant who still has to learn to make abstract distinctions, while the fully developed version of an adult would more like this:
 |
The abstract half is slightly elevated (abstract thinking has a proven extra value), getting slightly higherr the more abstract i.e. to the left (the more general a rule, the greater its predictive value), with the cubicles getting less in number and bigger (the abstracts encompass ever more). All according to the abstraction ladder.
Neurology → Sociology
There are two things in the above that exert a more direct influence on sociology. Firstly: those individual i.e. psychological, i.e. partly neurological aspects that get transported over the boundary when going from one individual to many.And secondly: the things that were stated about the workings of oxyticin and vasopressin, as neurotransmitters that work on turning a set of loose individuals into a group.
For the latter goes that the fact that they belong to neurological structures of the second level, they also must apply to a lot of evolutionary ancestors of humankind. A notable and noted example is that of group behaviour in apes, that has been quite extensively studied by several groups, leading e.g. to the popularly acclaimed Chimpanzee Politics by Frans de Waal
In humans it has been shown that the slightest form of external distinguishing sign between two groups of people, can lead to them flocking together and developing repulsive (literally and figuratively) behaviour towards each other
With human beings, there is an additional aspect to this process, that leads to more vulnaribility to it and greater severity of its consequences: the kind of communication called "language". The faculty of human language is that they can communicate much more precisely about the mental maps of reality they have made in their minds, thereby improving on these maps - the gaps in the one get filled in by the other, and the errors can get corrected. At least in some (or optimistically: many) cases.
But this has a flip side: systematic errors and misconceptions can also be easily spread around. One of them is the addiction to the higher abstractions, as explained in more detail in Abstraction ladder
In both cases, what language does is to provide an extra mode of feedback. In case this feedback is of the kind leading towards the equilibrium, i.e. the correct observation of reality, the extra feedback enhances the equilibrium. If the feedback leads away from the equilibrium, i.e. away from reality, i.e. towards concepts in the mind not related to reality, the feedback steers towards instability.
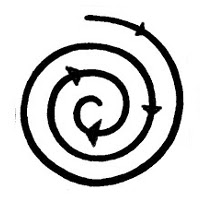 The
case of getting away from the equilibrium is in the theory of feedback
(called "cybernetics"
The
case of getting away from the equilibrium is in the theory of feedback
(called "cybernetics"
With this imagery in mind, it is not difficult to find all kinds of examples of the phenomena caused by the extra feedback through language, which examples go beyond the scope of this article, because some of them are quite to very widespread. And mentioning them is controversial to those that suffer from this state caused by external feedback.
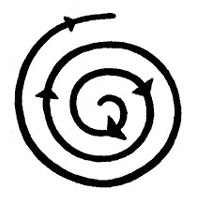 The
solution once in this state is of course to find a way to do the reverse
process, see the next illustration on the right. There are lots of ways to
endeavour on this path, which as far as the material aspects of live are
mainly represented by what is called the "sciences", for which in some
languages one has to specify "the natural sciences".
The
solution once in this state is of course to find a way to do the reverse
process, see the next illustration on the right. There are lots of ways to
endeavour on this path, which as far as the material aspects of live are
mainly represented by what is called the "sciences", for which in some
languages one has to specify "the natural sciences".For more general consumption, the guidelines provided by the already mentioned field of general semantics are at least as good as any of them.
Further developments
The logical next thing on the list is the description of how the cortex works together with emotion organs and brainstem - hopefully this will be done here sometime in the future. Undoubtedly it will lead to corrections to the presented models.For reading from this point on, one is referred to the already mentioned article on the hippocampus and the abstraction ladder
Some more neurology (on the cerebellum) here